

PRPF8 increases the aggressiveness of hepatocellular carcinoma by regulating FAK/AKT pathway via fibronectin 1 splicing
- PMID: 36609600
- PMCID: PMC9898568
- DOI: 10.1038/s12276-022-00917-7
PRPF8 increases the aggressiveness of hepatocellular carcinoma by regulating FAK/AKT pathway via fibronectin 1 splicing
Abstract
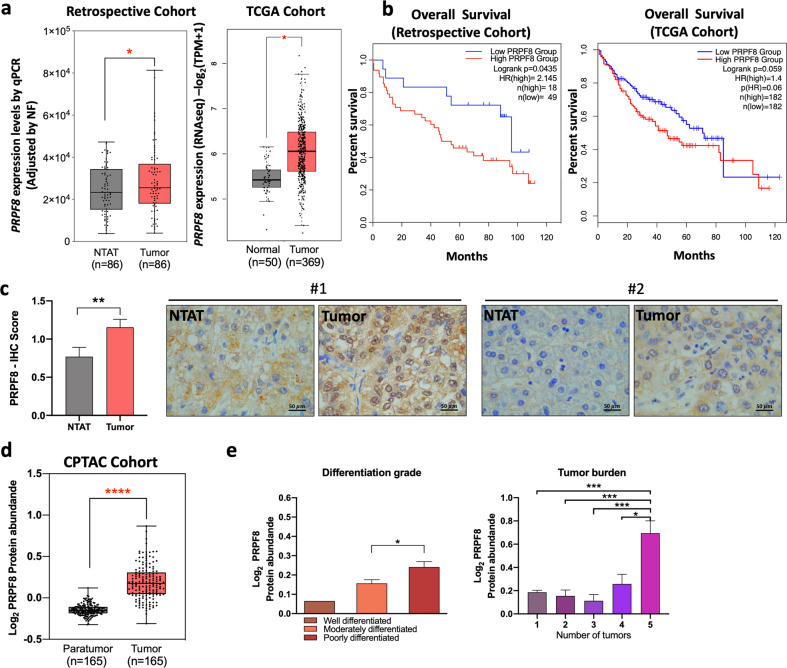
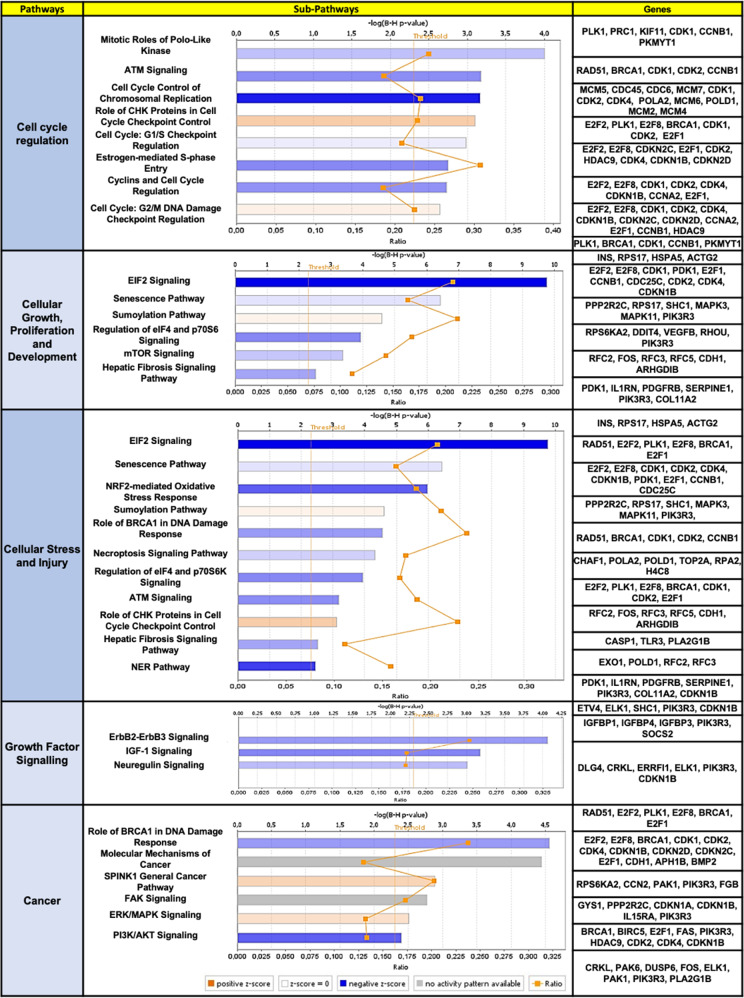
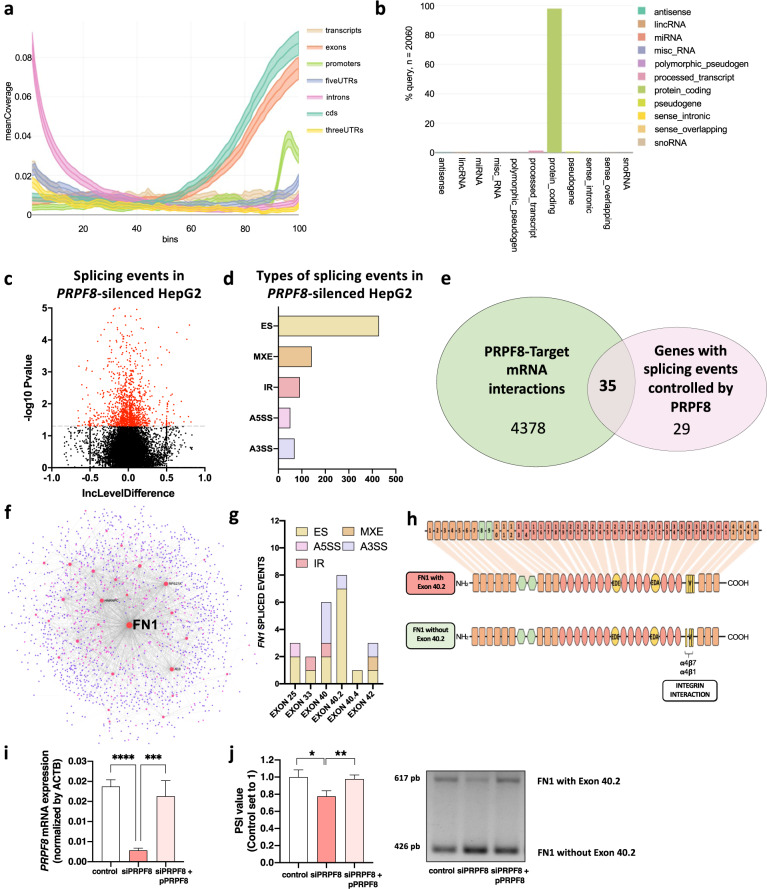
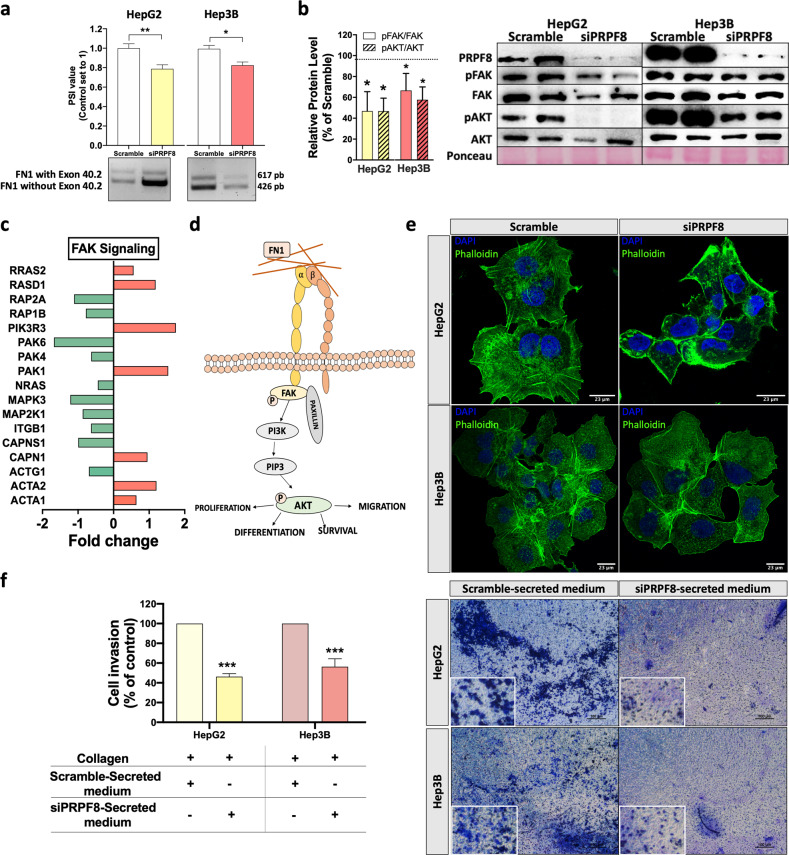
Hepatocellular carcinoma (HCC) pathogenesis is associated with alterations in splicing machinery components (spliceosome and splicing factors) and aberrant expression of oncogenic splice variants. We aimed to analyze the expression and potential role of the spliceosome component PRPF8 (pre-mRNA processing factor 8) in HCC. PRPF8 expression (mRNA/protein) was analyzed in a retrospective cohort of HCC patients (n = 172 HCC and nontumor tissues) and validated in two in silico cohorts (TCGA and CPTAC). PRPF8 expression was silenced in liver cancer cell lines and in xenograft tumors to understand the functional and mechanistic consequences. In silico RNAseq and CLIPseq data were also analyzed. Our results indicate that PRPF8 is overexpressed in HCC and associated with increased tumor aggressiveness (patient survival, etc.), expression of HCC-related splice variants, and modulation of critical genes implicated in cancer-related pathways. PRPF8 silencing ameliorated aggressiveness in vitro and decreased tumor growth in vivo. Analysis of in silico CLIPseq data in HepG2 cells demonstrated that PRPF8 binds preferentially to exons of protein-coding genes, and RNAseq analysis showed that PRPF8 silencing alters splicing events in multiple genes. Integrated and in vitro analyses revealed that PRPF8 silencing modulates fibronectin (FN1) splicing, promoting the exclusion of exon 40.2, which is paramount for binding to integrins. Consistent with this finding, PRPF8 silencing reduced FAK/AKT phosphorylation and blunted stress fiber formation. Indeed, HepG2 and Hep3B cells exhibited a lower invasive capacity in membranes treated with conditioned medium from PRPF8-silenced cells compared to medium from scramble-treated cells. This study demonstrates that PRPF8 is overexpressed and associated with aggressiveness in HCC and plays important roles in hepatocarcinogenesis by altering FN1 splicing, FAK/AKT activation and stress fiber formation.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
