

Ubiquilin-2 regulates pathological alpha-synuclein
- PMID: 36609661
- PMCID: PMC9823102
- DOI: 10.1038/s41598-022-26899-0
Ubiquilin-2 regulates pathological alpha-synuclein
Abstract
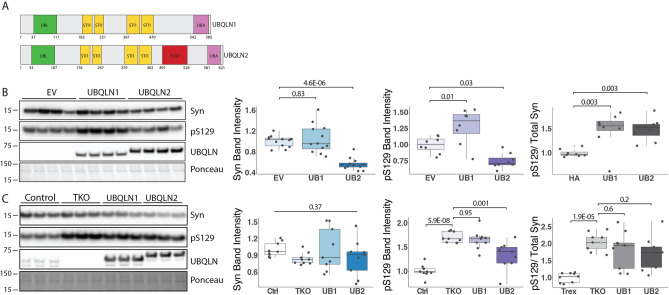
The key protein implicated in Parkinson's disease and other synucleinopathies is α-synuclein, and a post-translationally modified form of the protein, phosphorylated at serine 129 (pS129), is a principal component in Lewy bodies, a pathological hallmark of PD. While altered proteostasis has been implicated in the etiology of Parkinson's disease, we still have a limited understanding of how α-synuclein is regulated in the nervous system. The protein quality control protein Ubiquilin-2 (UBQLN2) is known to accumulate in synucleinopathies, but whether it directly regulates α-synuclein is unknown. Using cellular and mouse models, we find that UBQLN2 decreases levels of α-synuclein, including the pS129 phosphorylated isoform. Pharmacological inhibition of the proteasome revealed that, while α-synuclein may be cleared by parallel and redundant quality control pathways, UBQLN2 preferentially targets pS129 for proteasomal degradation. Moreover, in brain tissue from human PD and transgenic mice expressing pathogenic α-synuclein (A53T), native UBQLN2 becomes more insoluble. Collectively, our studies support a role for UBQLN2 in directly regulating pathological forms of α-synuclein and indicate that UBQLN2 dysregulation in disease may contribute to α-synuclein-mediated toxicity.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
