

Persistent nociceptor hyperactivity as a painful evolutionary adaptation
- PMID: 36610893
- PMCID: PMC9974896
- DOI: 10.1016/j.tins.2022.12.007
Persistent nociceptor hyperactivity as a painful evolutionary adaptation
Abstract
Chronic pain caused by injury or disease of the nervous system (neuropathic pain) has been linked to persistent electrical hyperactivity of the sensory neurons (nociceptors) specialized to detect damaging stimuli and/or inflammation. This pain and hyperactivity are considered maladaptive because both can persist long after injured tissues have healed and inflammation has resolved. While the assumption of maladaptiveness is appropriate in many diseases, accumulating evidence from diverse species, including humans, challenges the assumption that neuropathic pain and persistent nociceptor hyperactivity are always maladaptive. We review studies indicating that persistent nociceptor hyperactivity has undergone evolutionary selection in widespread, albeit selected, animal groups as a physiological response that can increase survival long after bodily injury, using both highly conserved and divergent underlying mechanisms.
Keywords: Aplysia; cephalopod; chronic pain; human; primary afferent neuron; spontaneous activity.
Copyright © 2022 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests T.J.P. is a co-founder of 4E Therapeutics, Doloromics, PARMedics, and NuvoNuro; he serves on the Board of Directors of 4E Therapeutics and Doloromics; he is an inventor on patents related to MNK inhibition for the treatment of pain. E.S.S. receives funding from AstraZeneca and GlaxoSmithKline. E.T.W., R.J.C., and G.G.N. declare no competing interests.
Figures
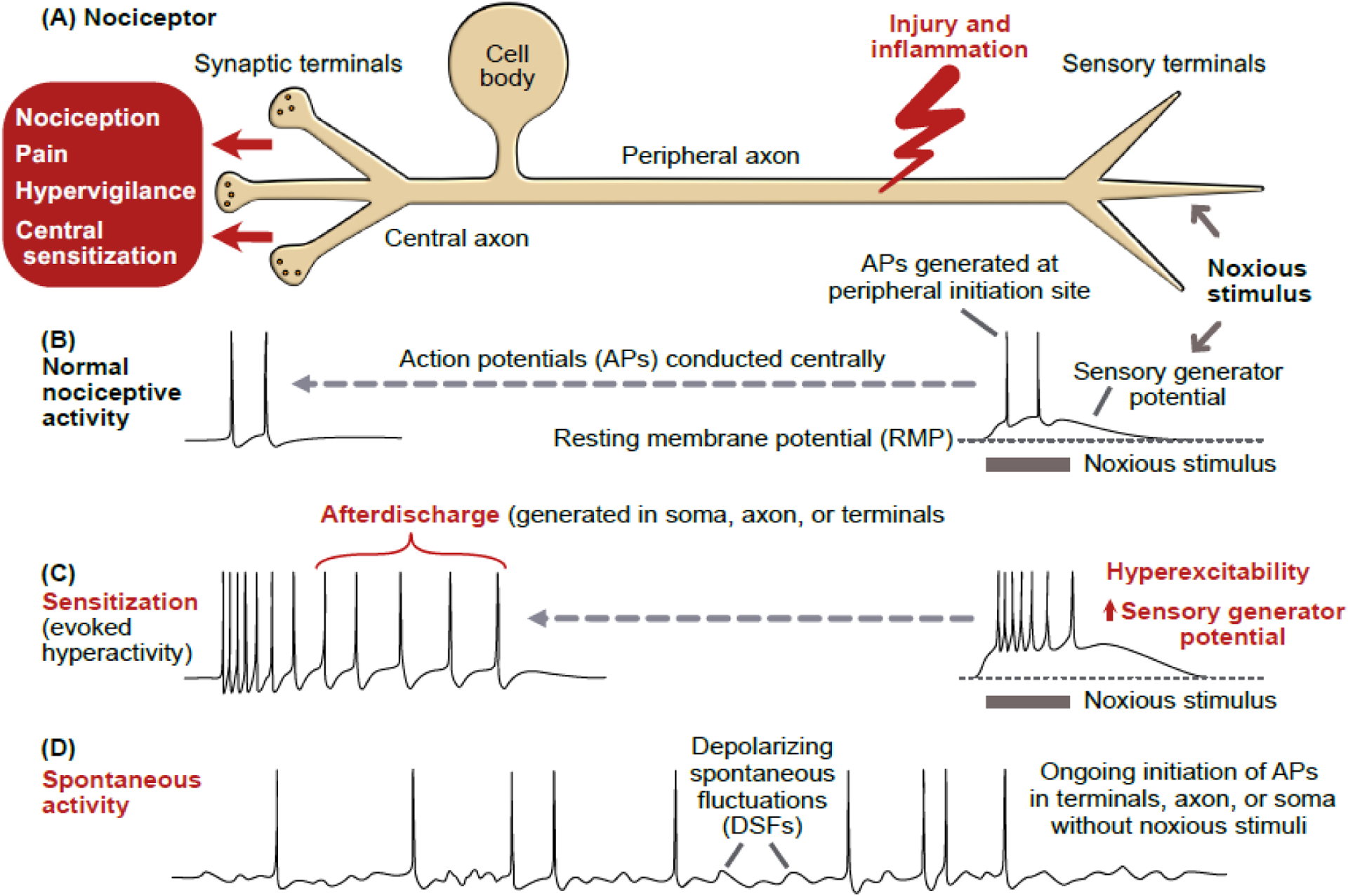
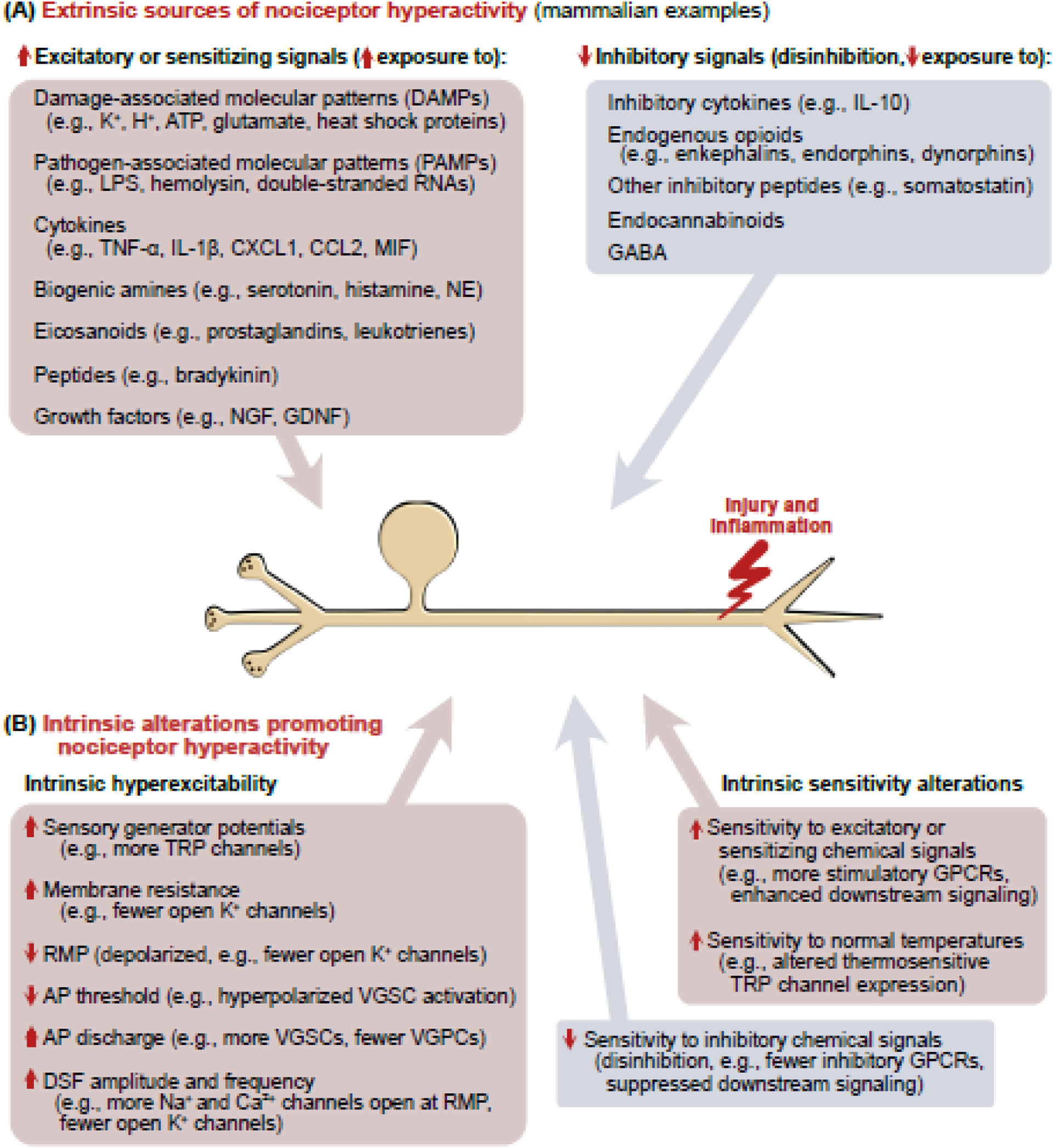
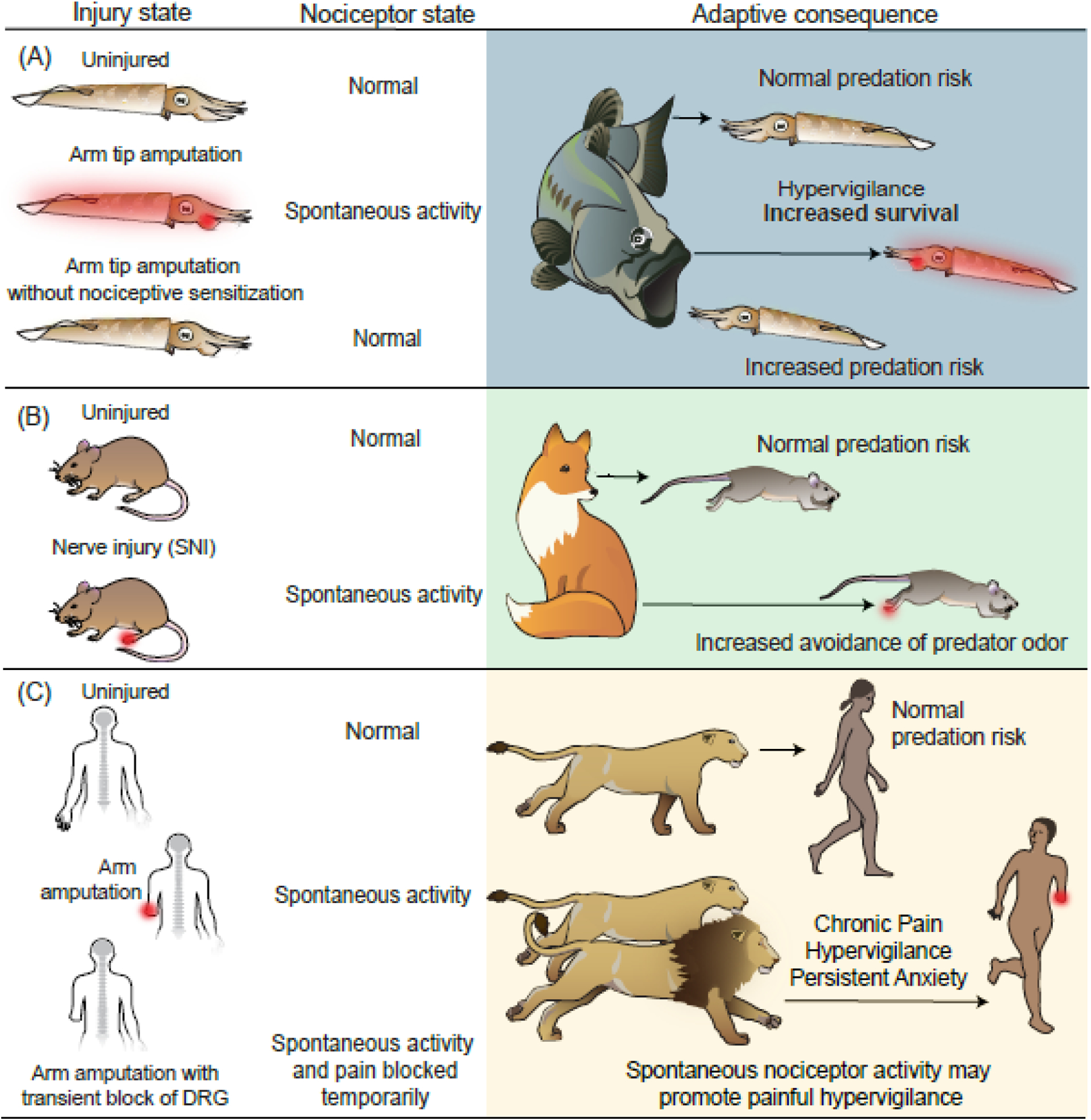
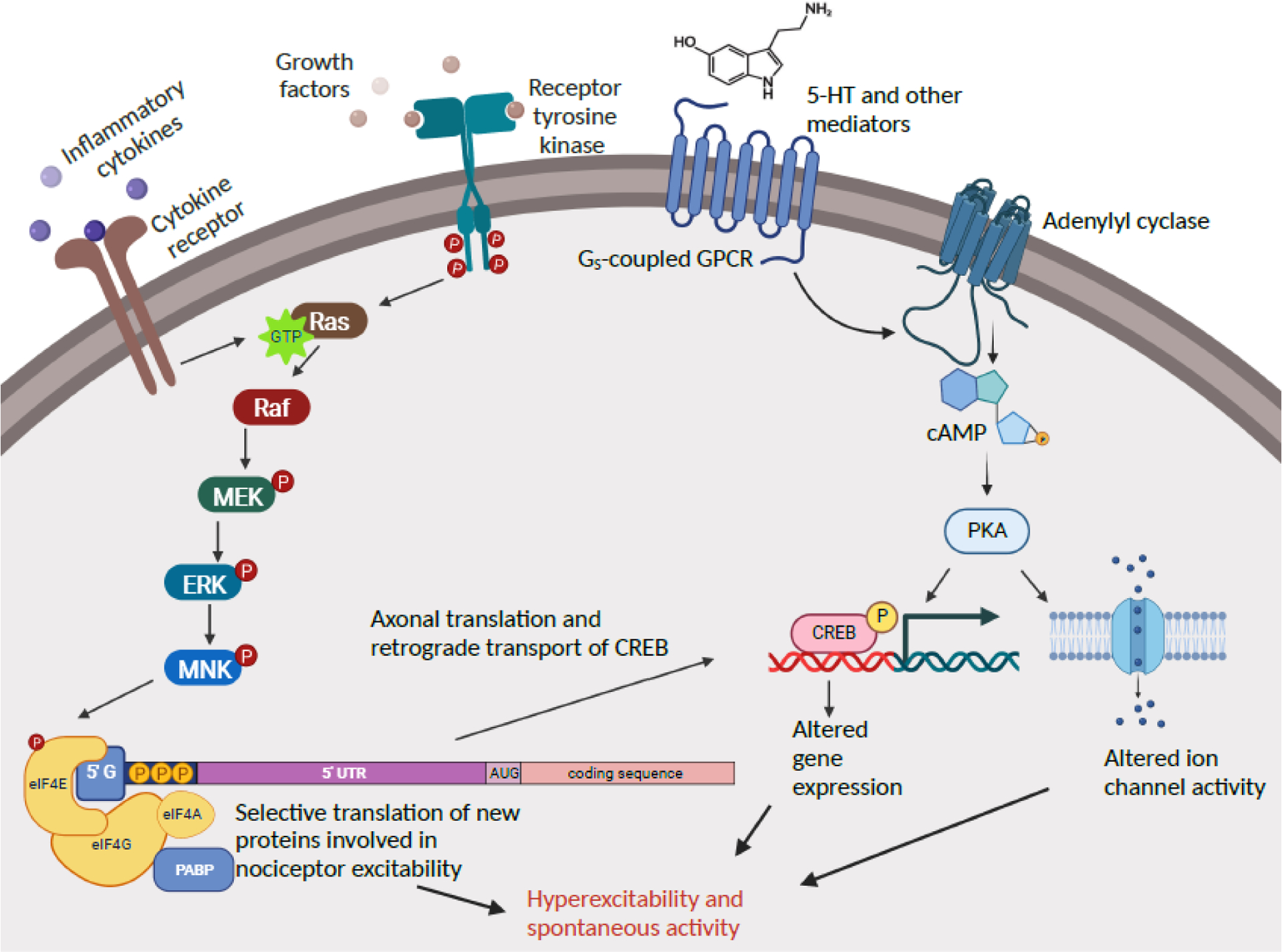
References
-
- Williams GC and Nesse RM (1991) The dawn of Darwinian medicine. Q Rev Biol 66, 1–22 - PubMed
-
- Finnerup NB et al. (2021) Neuropathic Pain: From Mechanisms to Treatment. Physiol Rev 101, 259–301 - PubMed
-
- Sommer C et al. (2018) Inflammation in the pathophysiology of neuropathic pain. Pain 159, 595–602 - PubMed
-
- Walters ET (2021) Nociceptors and chronic pain. In Oxford Research Encyclopedia of Neuroscience (Murray S. Oxford University Press
-
- Roza C and Bernal L (2022) Electrophysiological characterization of ectopic spontaneous discharge in axotomized and intact fibers upon nerve transection: a role in spontaneous pain. Pflugers Arch 474, 387–396 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
