

Intra-colony venom diversity contributes to maintaining eusociality in a cooperatively breeding ant
- PMID: 36617555
- PMCID: PMC9827630
- DOI: 10.1186/s12915-022-01507-9
Intra-colony venom diversity contributes to maintaining eusociality in a cooperatively breeding ant
Abstract
Background: Eusociality is widely considered to evolve through kin selection, where the reproductive success of an individual's close relative is favored at the expense of its own. High genetic relatedness is thus considered a prerequisite for eusociality. While ants are textbook examples of eusocial animals, not all ants form colonies of closely related individuals. One such example is the ectatommine ant Rhytidoponera metallica, which predominantly forms queen-less colonies that have such a low intra-colony relatedness that they have been proposed to represent a transient, unstable form of eusociality. However, R. metallica is among the most abundant and widespread ants on the Australian continent. This apparent contradiction provides an example of how inclusive fitness may not by itself explain the maintenance of eusociality and raises the question of what other selective advantages maintain the eusocial lifestyle of this species.
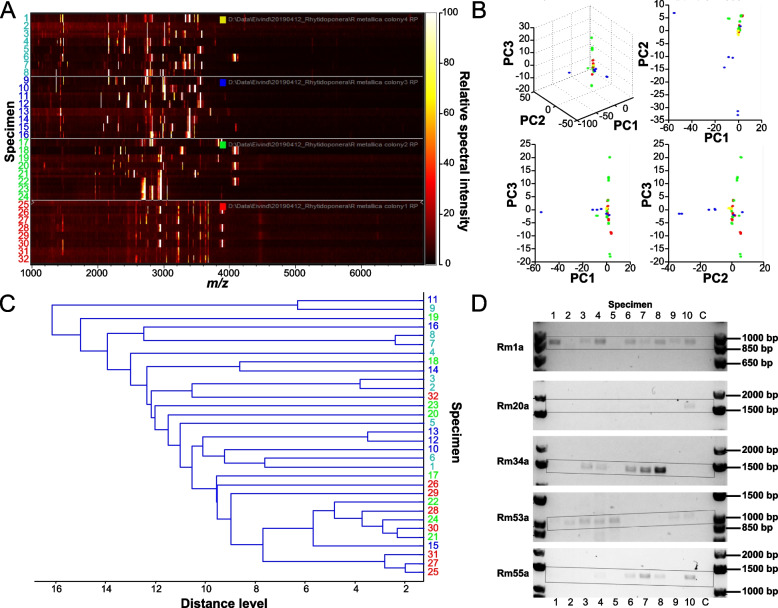
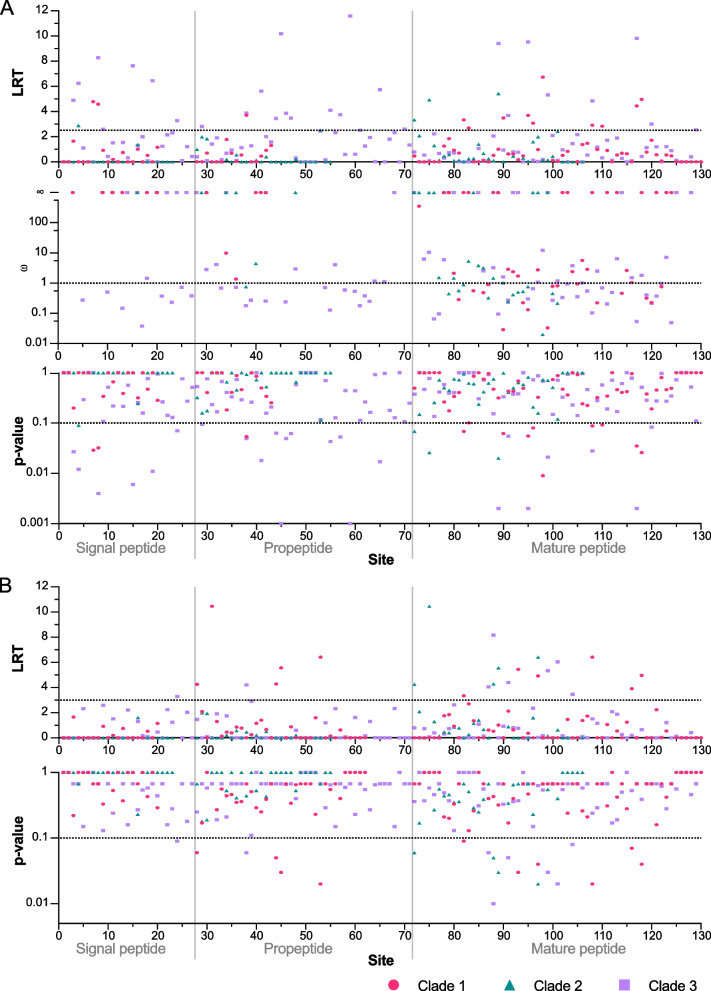
Results: We provide a comprehensive portrait of the venom of R. metallica and show that the colony-wide venom consists of an exceptionally high diversity of functionally distinct toxins for an ant. These toxins have evolved under strong positive selection, which is normally expected to reduce genetic variance. Yet, R. metallica exhibits remarkable intra-colony variation, with workers sharing only a relatively small proportion of toxins in their venoms. This variation is not due to the presence of chemical castes, but has a genetic foundation that is at least in part explained by toxin allelic diversity.
Conclusions: Taken together, our results suggest that the toxin diversity contained in R. metallica colonies may be maintained by a form of group selection that selects for colonies that can exploit more resources and defend against a wider range of predators. We propose that increased intra-colony genetic variance resulting from low kinship may itself provide a selective advantage in the form of an expanded pharmacological venom repertoire. These findings provide an example of how group selection on adaptive phenotypes may contribute to maintaining eusociality where a prerequisite for kin selection is diminished.
Keywords: Eusocial; Formicidae; Group selection; Kin selection; Peptide; Toxin.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
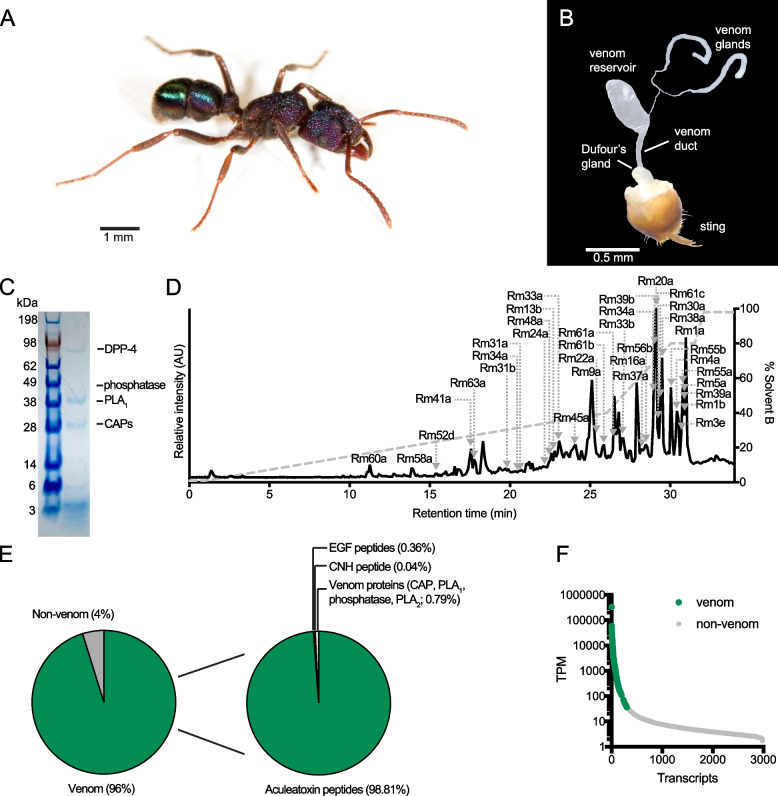

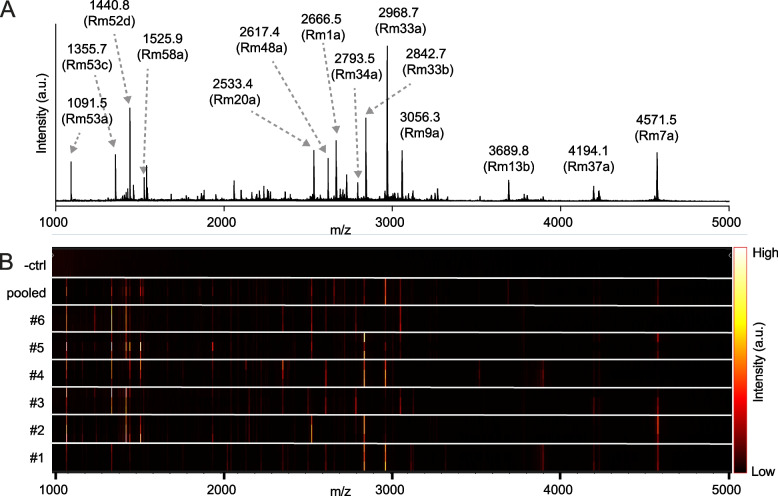
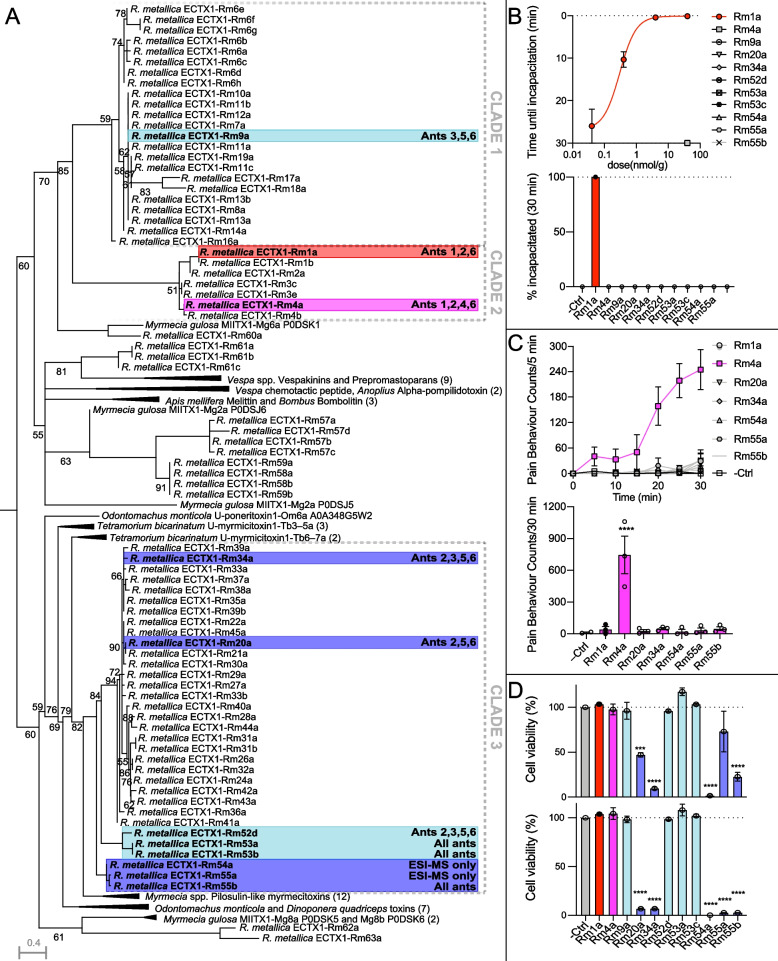
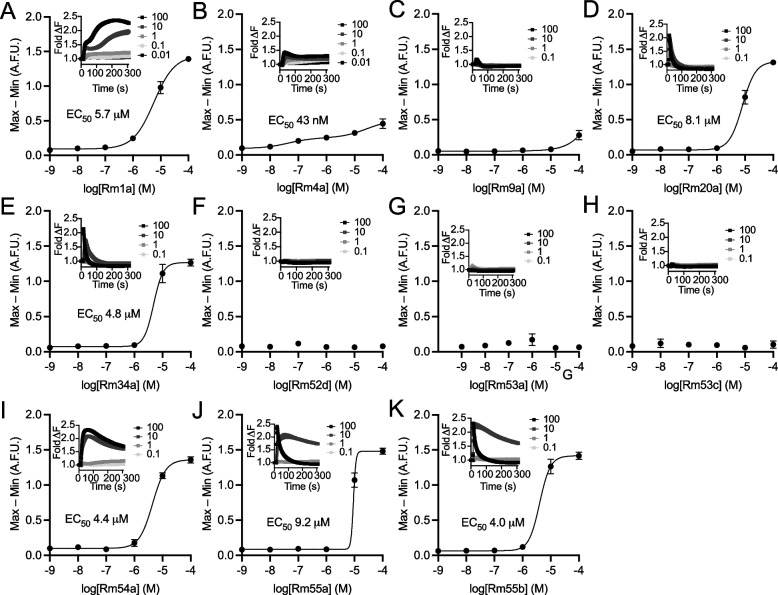
References
-
- Hughes WOH, Boomsma JJ. Genetic diversity and disease resistance in leaf-cutting ant societies. Evolution. 2004;58:1251–1260. - PubMed
-
- Haskins CP, Haskins EF. Situation and location-specific factors in the compatibility response in Rhytidoponera metallica (Hymenoptera: Formicidae: Ponerinae) Psyche. 1983;90:163–174. doi: 10.1155/1983/67615. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
