

Progranulin loss results in sex-dependent dysregulation of the peripheral and central immune system
- PMID: 36618392
- PMCID: PMC9814971
- DOI: 10.3389/fimmu.2022.1056417
Progranulin loss results in sex-dependent dysregulation of the peripheral and central immune system
Abstract
Introduction: Progranulin (PGRN) is a secreted glycoprotein, the expression of which is linked to several neurodegenerative diseases. Although its specific function is still unclear, several studies have linked it with lysosomal functions and immune system regulation. Here, we have explored the role of PGRN in peripheral and central immune system homeostasis by investigating the consequences of PGRN deficiency on adaptive and innate immune cell populations.
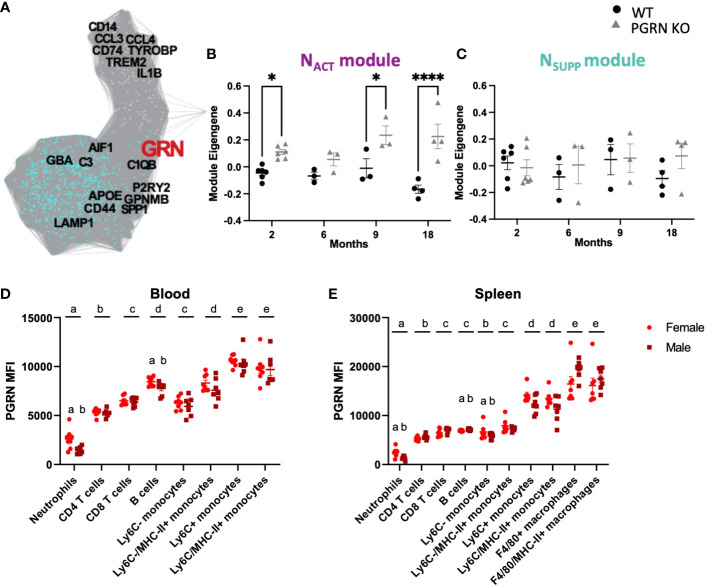
Methods: First, we used gene co-expression network analysis of published data to test the hypothesis that Grn has a critical role in regulating the activation status of immune cell populations in both central and peripheral compartments. To investigate the extent to which PGRN-deficiency resulted in immune dysregulation, we performed deep immunophenotyping by flow cytometry of 19-24-month old male and female Grn-deficient mice (PGRN KO) and littermate Grn-sufficient controls (WT).
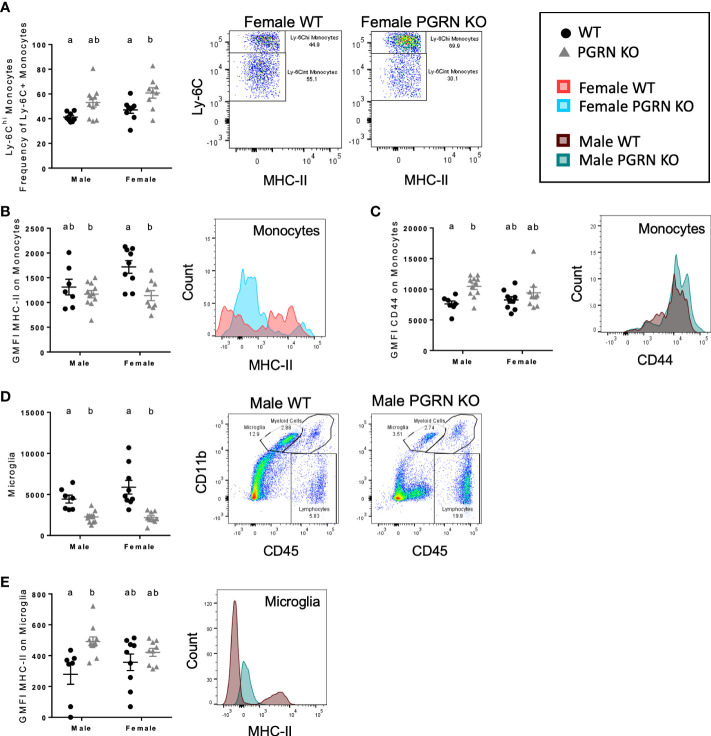
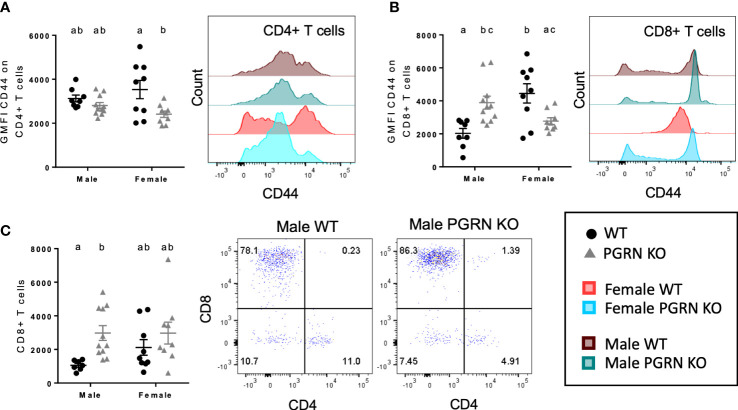
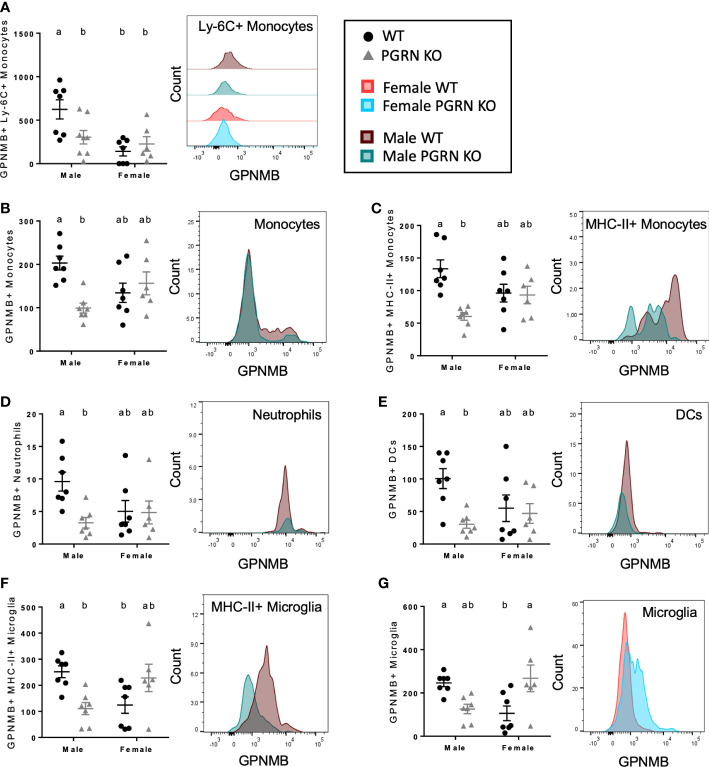
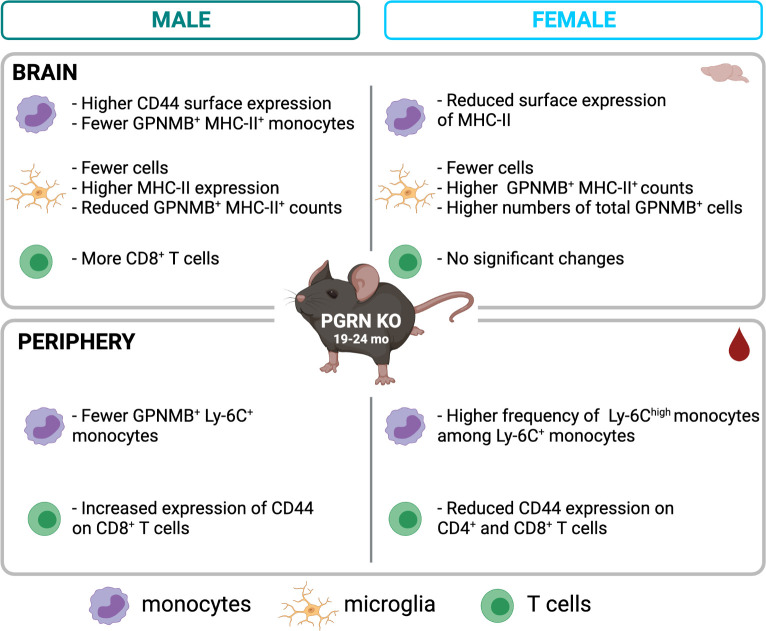
Results: Male PGRN KO mice exhibited a lower abundance of microglial cells with higher MHC-II expression, increased CD44 expression on monocytes in the brain, and more CNS-associated CD8+ T cells compared to WT mice. Furthermore, we observed an increase in CD44 on CD8+ T cells in the peripheral blood. Female PGRN KO mice also had fewer microglia compared to WT mice, and we also observed reduced expression of MHC-II on brain monocytes. Additionally, we found an increase in Ly-6Chigh monocyte frequency and decreased CD44 expression on CD8+ and CD4+ T cells in PGRN KO female blood. Given that Gpnmb, which encodes for the lysosomal protein Glycoprotein non-metastatic melanoma protein B, has been reported to be upregulated in PGRN KO mice, we investigated changes in GPNMB protein expression associated with PGRN deficits and found that GPNMB is modulated in myeloid cells in a sex-specific manner.
Discussion: Our data suggest that PGRN and GPNMB jointly regulate the peripheral and the central immune system in a sex-specific manner; thus, understanding their associated mechanisms could pave the way for developing new neuroprotective strategies to modulate central and peripheral inflammation to lower risk for neurodegenerative diseases and possibly delay or halt progression.
Keywords: GPNMB; T cells; microglia; monocytes; neurodegenaration; peripheral-brain crosstalk; progranulin.
Copyright © 2022 Houser, Uriarte Huarte, Wallings, Keating, MacPherson, Herrick, Kannarkat, Kelly, Chang, Varvel, Rexach and Tansey.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous