

RanBP9 controls the oligomeric state of CTLH complex assemblies
- PMID: 36621627
- PMCID: PMC9932110
- DOI: 10.1016/j.jbc.2023.102869
RanBP9 controls the oligomeric state of CTLH complex assemblies
Abstract
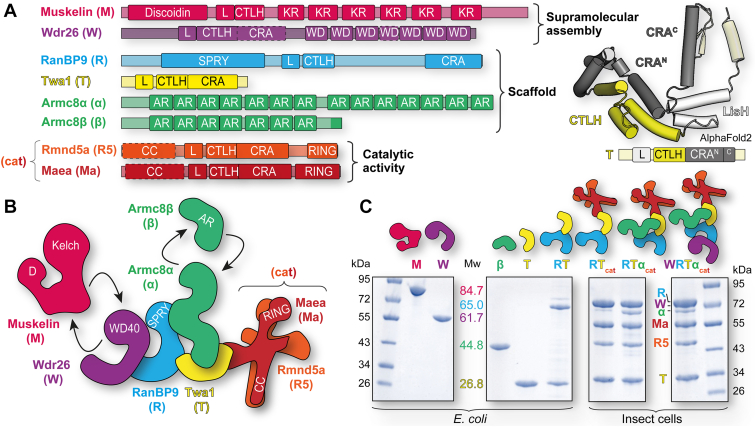
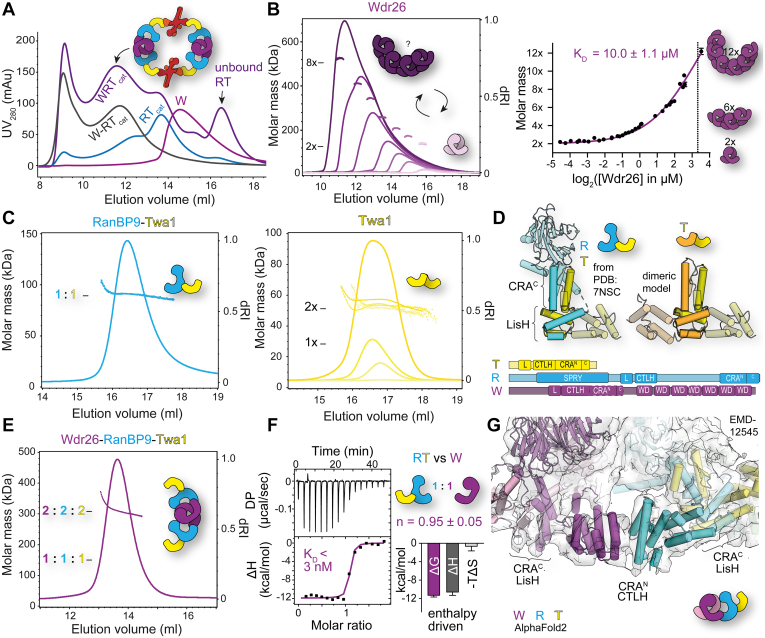
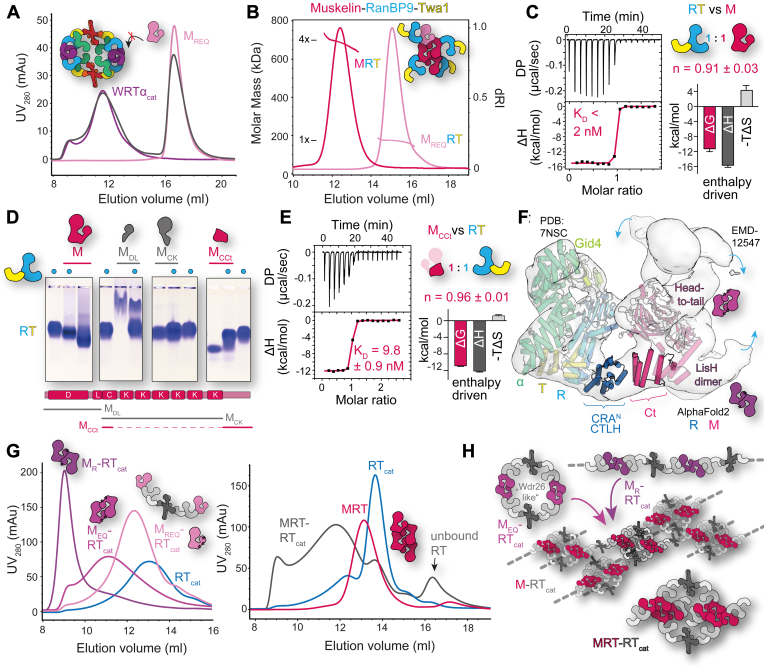
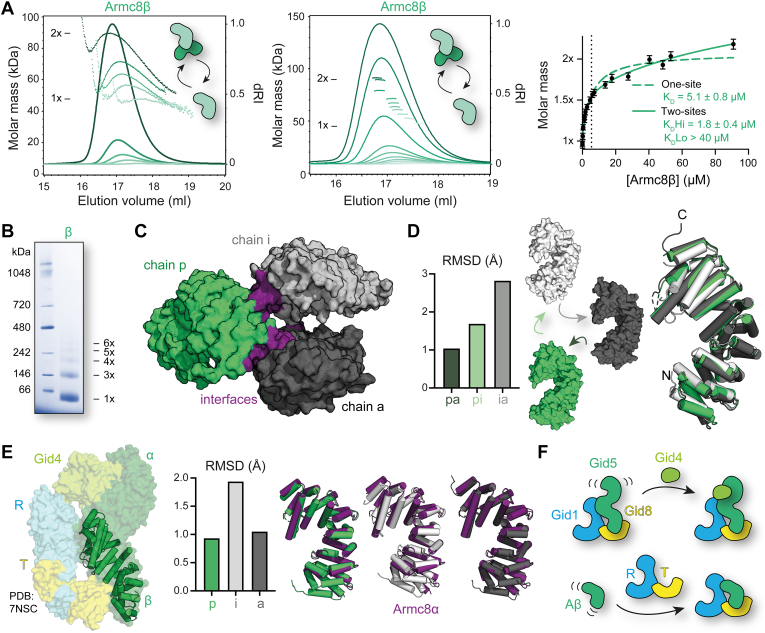
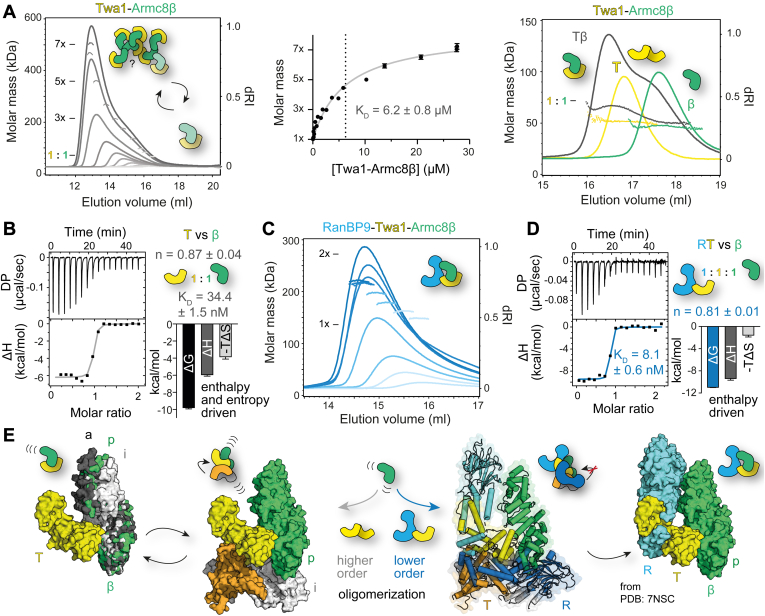
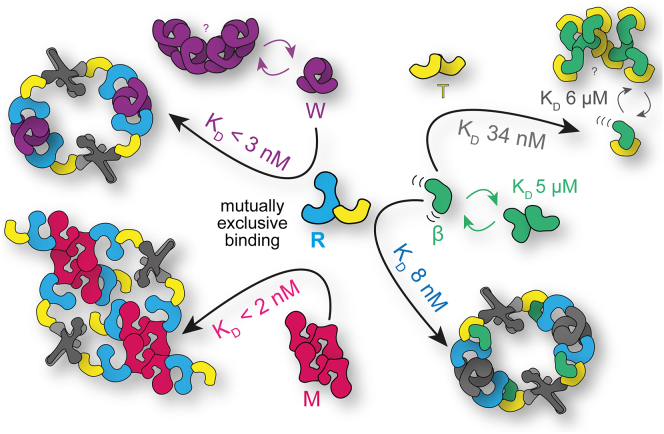
The CTLH (C-terminal to lissencephaly-1 homology motif) complex is a multisubunit RING E3 ligase with poorly defined substrate specificity and flexible subunit composition. Two key subunits, muskelin and Wdr26, specify two alternative CTLH complexes that differ in quaternary structure, thereby allowing the E3 ligase to presumably target different substrates. With the aid of different biophysical and biochemical techniques, we characterized CTLH complex assembly pathways, focusing not only on Wdr26 and muskelin but also on RanBP9, Twa1, and Armc8β subunits, which are critical to establish the scaffold of this E3 ligase. We demonstrate that the ability of muskelin to tetramerize and the assembly of Wdr26 into dimers define mutually exclusive oligomerization modules that compete with nanomolar affinity for RanBP9 binding. The remaining scaffolding subunits, Armc8β and Twa1, strongly interact with each other and with RanBP9, again with nanomolar affinity. Our data demonstrate that RanBP9 organizes subunit assembly and prevents higher order oligomerization of dimeric Wdr26 and the Armc8β-Twa1 heterodimer through its tight binding. Combined, our studies define alternative assembly pathways of the CTLH complex and elucidate the role of RanBP9 in governing differential oligomeric assemblies, thereby advancing our mechanistic understanding of CTLH complex architectures.
Keywords: GID–CTLH complex; ITC; RING E3 ligase; SEC–MALS; X-ray crystallography.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Deshaies R.J., Joazeiro C.A.P. RING domain E3 ubiquitin ligases. Annu. Rev. Biochem. 2009;78:399–434. - PubMed
-
- Hershko A., Ciechanover A. The ubiquitin system. Annu. Rev. Biochem. 1998;67:425–479. - PubMed
-
- Zheng N., Shabek N. Ubiquitin ligases: structure, function, and regulation. Annu. Rev. Biochem. 2017;86:129–157. - PubMed
-
- Kobayashi N., Yang J., Ueda A., Suzuki T., Tomaru K., Takeno M., et al. RanBPM, Muskelin, p48EMLP, p44CTLH, and the armadillo-repeat proteins ARMC8alpha and ARMC8beta are components of the CTLH complex. Gene. 2007;396:236–247. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
