

Membrane skeleton hyperstability due to a novel alternatively spliced 4.1R can account for ellipsoidal camelid red cells with decreased deformability
- PMID: 36621628
- PMCID: PMC9926112
- DOI: 10.1016/j.jbc.2023.102877
Membrane skeleton hyperstability due to a novel alternatively spliced 4.1R can account for ellipsoidal camelid red cells with decreased deformability
Abstract
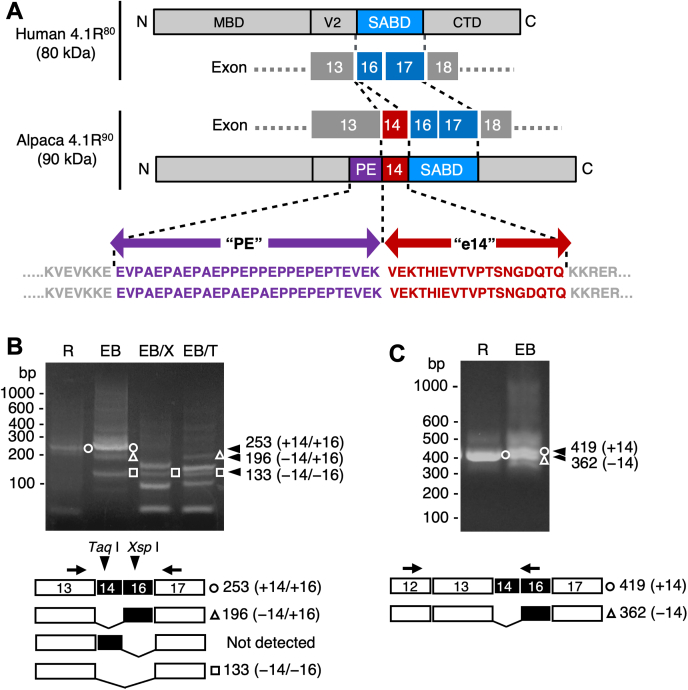
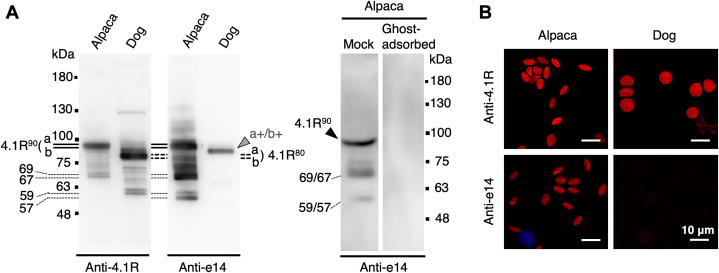
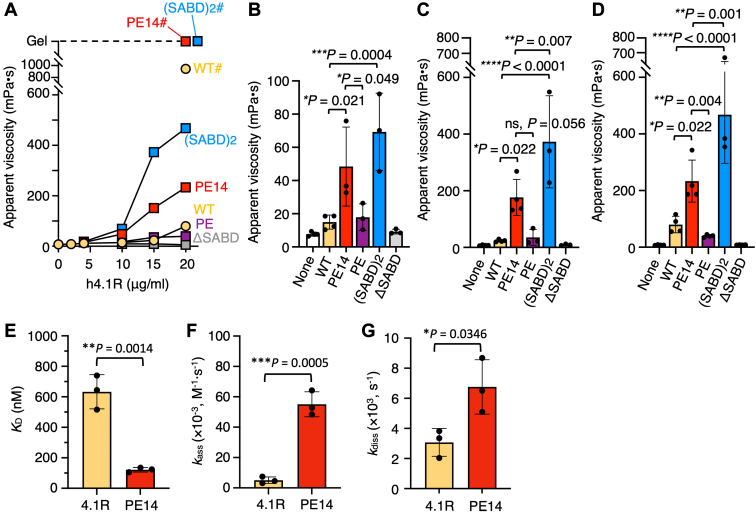
The red blood cells (RBCs) of vertebrates have evolved into two basic shapes, with nucleated nonmammalian RBCs having a biconvex ellipsoidal shape and anuclear mammalian RBCs having a biconcave disk shape. In contrast, camelid RBCs are flat ellipsoids with reduced membrane deformability, suggesting altered membrane skeletal organization. However, the mechanisms responsible for their elliptocytic shape and reduced deformability have not been determined. We here showed that in alpaca RBCs, protein 4.1R, a major component of the membrane skeleton, contains an alternatively spliced exon 14-derived cassette (e14) not observed in the highly conserved 80 kDa 4.1R of other highly deformable biconcave mammalian RBCs. The inclusion of this exon, along with the preceding unordered proline- and glutamic acid-rich peptide (PE), results in a larger and unique 90 kDa camelid 4.1R. Human 4.1R containing e14 and PE, but not PE alone, showed markedly increased ability to form a spectrin-actin-4.1R ternary complex in viscosity assays. A similar facilitated ternary complex was formed by human 4.1R possessing a duplication of the spectrin-actin-binding domain, one of the mutations known to cause human hereditary elliptocytosis. The e14- and PE-containing mutant also exhibited an increased binding affinity to β-spectrin compared with WT 4.1R. Taken together, these findings indicate that 4.1R protein with the e14 cassette results in the formation and maintenance of a hyperstable membrane skeleton, resulting in rigid red ellipsoidal cells in camelid species, and suggest that membrane structure is evolutionarily regulated by alternative splicing of exons in the 4.1R gene.
Keywords: alternative splicing; camelid; elliptocyte; erythrocyte; hereditary elliptocytosis; membrane protein; membrane structure; plasma membrane; protein 4.1R; spectrin.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures


References
-
- Lazarides E. From genes to structural morphogenesis: the genesis and epigenesis of a red blood cell. Cell. 1987;51:345–356. - PubMed
-
- Lazarides E., Woods C. Biogenesis of the red blood cell membrane-skeleton and the control of erythroid morphogenesis. Annu. Rev. Cell Biol. 1989;5:427–452. - PubMed
-
- Fowler V.M. Chapter two–the human erythrocyte plasma membrane: a Rosetta stone for decoding membrane-cytoskeleton structure. Curr. Top. Membr. 2013;72:39–88. - PubMed
-
- Gallagher P.G. Hereditary elliptocytosis: spectrin and protein 4.1R. Sem Hematol. 2004;41:142–164. - PubMed
-
- Gallagher P.G. In: Hematology: Basic Principles and Practice. 7th Ed. Hoffman R., Benz E.J. Jr., Silberstein L.E., Heslop H.E., Weitz J.I., Anastasi J., et al., editors. Elsevier; Philadelphia, PA: 2017. Red blood cell membrane disorder.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
