

Aspergillus fumigatus Drives Tissue Damage via Iterative Assaults upon Mucosal Integrity and Immune Homeostasis
- PMID: 36625602
- PMCID: PMC9933693
- DOI: 10.1128/iai.00333-22
Aspergillus fumigatus Drives Tissue Damage via Iterative Assaults upon Mucosal Integrity and Immune Homeostasis
Abstract
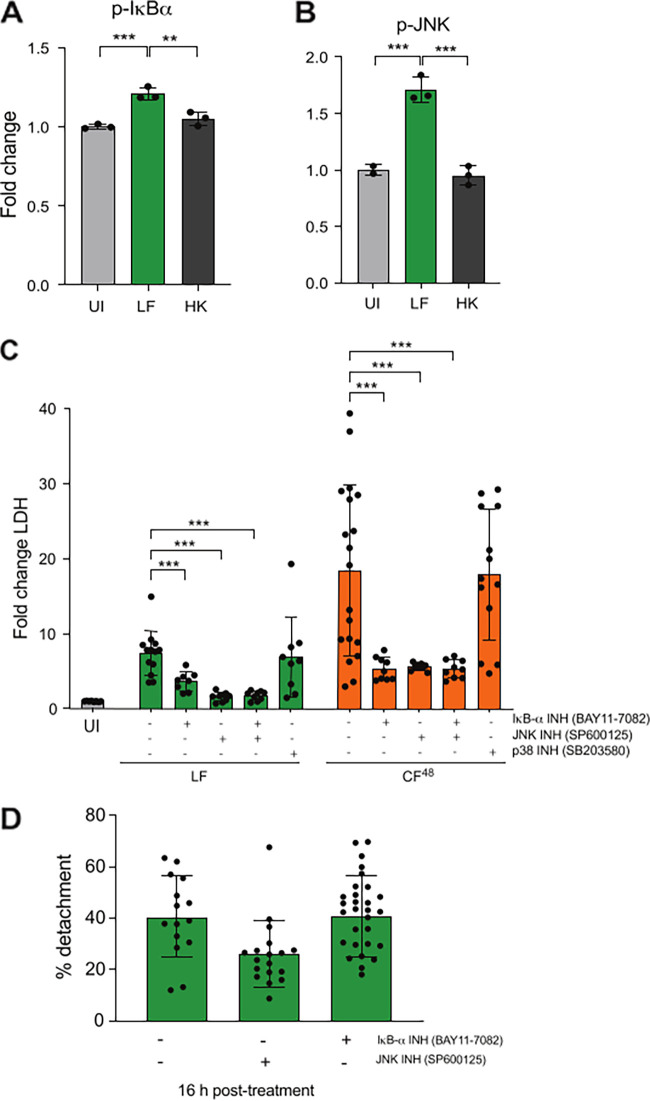
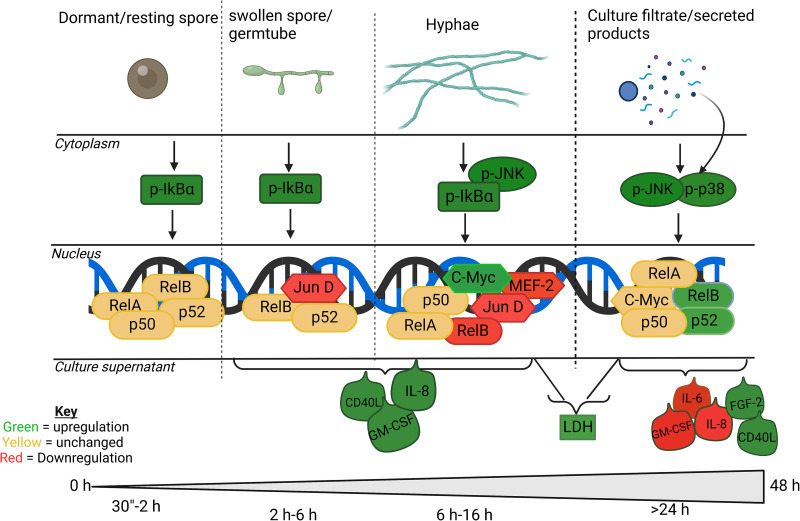
The human lung is constantly exposed to Aspergillus fumigatus spores, the most prevalent worldwide cause of fungal respiratory disease. Pulmonary tissue damage is a unifying feature of Aspergillus-related diseases; however, the mechanistic basis of damage is not understood. In the lungs of susceptible hosts, A. fumigatus undergoes an obligatory morphological switch involving spore germination and hyphal growth. We modeled A. fumigatus infection in cultured A549 human pneumocytes, capturing the phosphoactivation status of five host signaling pathways, nuclear translocation and DNA binding of eight host transcription factors, and expression of nine host response proteins over six time points encompassing exposures to live fungus and the secretome thereof. The resulting data set, comprised of more than 1,000 data points, reveals that pneumocytes mount differential responses to A. fumigatus spores, hyphae, and soluble secreted products via the NF-κB, JNK, and JNK + p38 pathways, respectively. Importantly, via selective degradation of host proinflammatory (IL-6 and IL-8) cytokines and growth factors (FGF-2), fungal secreted products reorchestrate the host response to fungal challenge as well as driving multiparameter epithelial damage, culminating in cytolysis. Dysregulation of NF-κB signaling, involving sequential stimulation of canonical and noncanonical signaling, was identified as a significant feature of host damage both in vitro and in a mouse model of invasive aspergillosis. Our data demonstrate that composite tissue damage results from iterative (repeated) exposures to different fungal morphotypes and secreted products and suggest that modulation of host responses to fungal challenge might represent a unified strategy for therapeutic control of pathologically distinct types of Aspergillus-related disease.
Keywords: Aspergillus fumigatus; fungal infection; lung infection; virulence.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
-
- Bertuzzi M, Schrettl M, Alcazar-Fuoli L, Cairns TC, Muñoz A, Walker LA, Herbst S, Safari M, Cheverton AM, Chen D, Liu H, Saijo S, Fedorova ND, Armstrong-James D, Munro CA, Read ND, Filler SG, Espeso EA, Nierman WC, Haas H, Bignell EM. 2014. The pH-responsive PacC transcription factor of Aspergillus fumigatus governs epithelial entry and tissue invasion during pulmonary aspergillosis. PLoS Pathog 10:e1004413. 10.1371/journal.ppat.1004413. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MR/S001824/1/MRC_/Medical Research Council/United Kingdom
- 214229_Z_18_Z/WT_/Wellcome Trust/United Kingdom
- MR/V031287/1/MRC_/Medical Research Council/United Kingdom
- MR/M02010X/1/MRC_/Medical Research Council/United Kingdom
- IS-BRC-1215-20006/DH_/Department of Health/United Kingdom
- MR/S001824/2/MRC_/Medical Research Council/United Kingdom
- MR/L000822/1/MRC_/Medical Research Council/United Kingdom
- MR/V033417/1/MRC_/Medical Research Council/United Kingdom
- BB/V017004/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 097377/Z/11/Z/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Medical
Research Materials
