

Resolving colistin resistance and heteroresistance in Enterobacter species
- PMID: 36627272
- PMCID: PMC9832134
- DOI: 10.1038/s41467-022-35717-0
Resolving colistin resistance and heteroresistance in Enterobacter species
Abstract
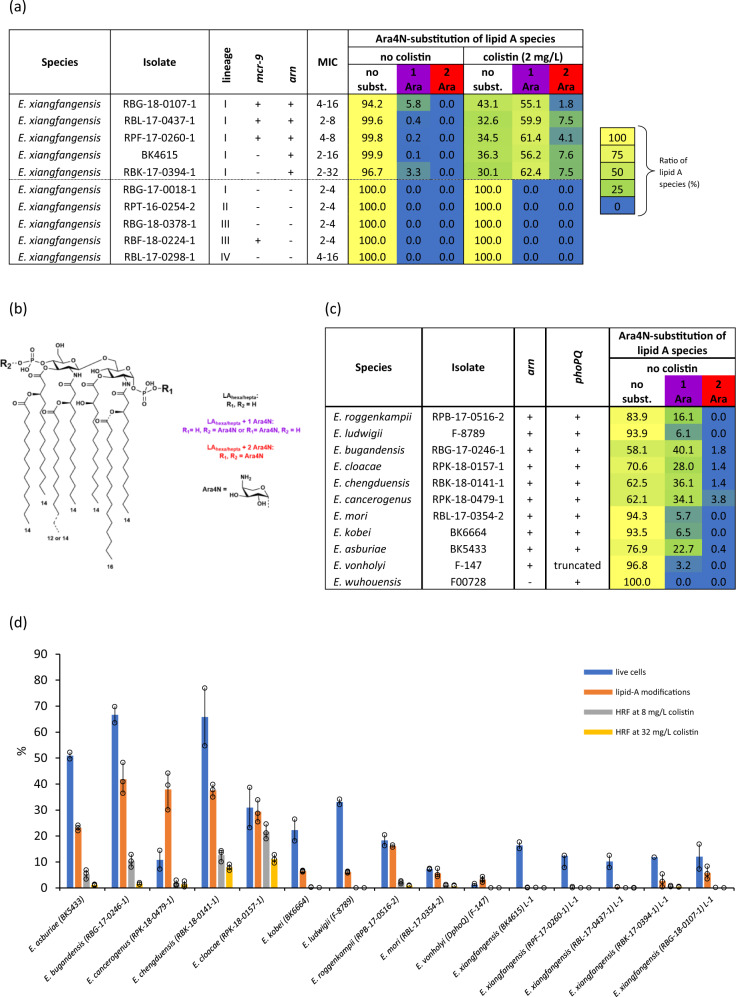
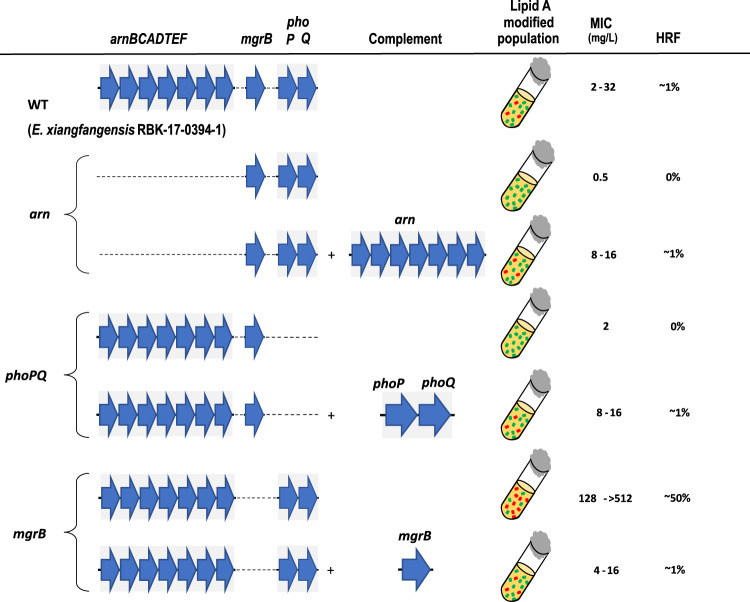
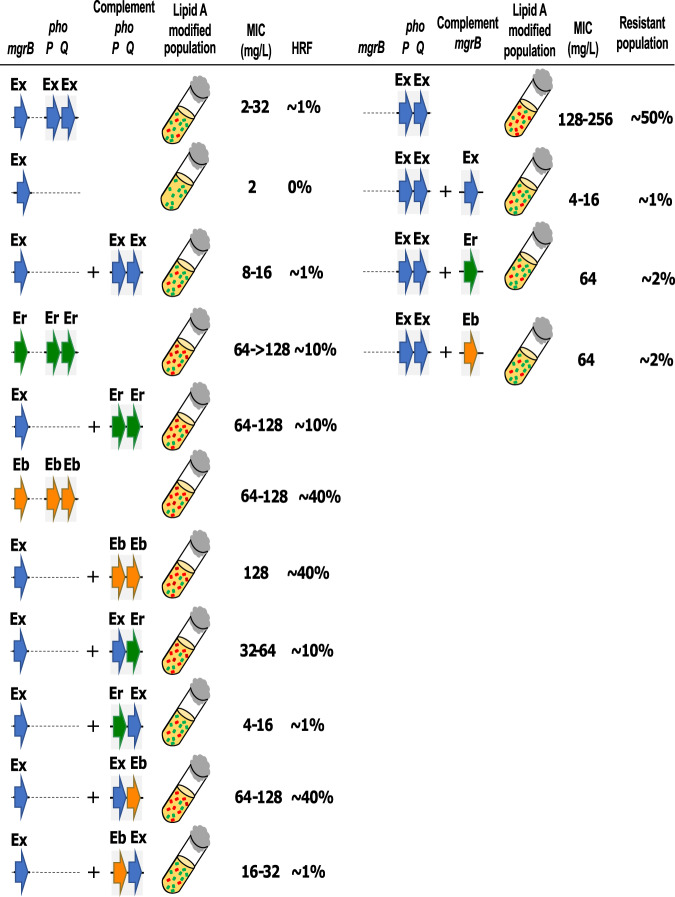
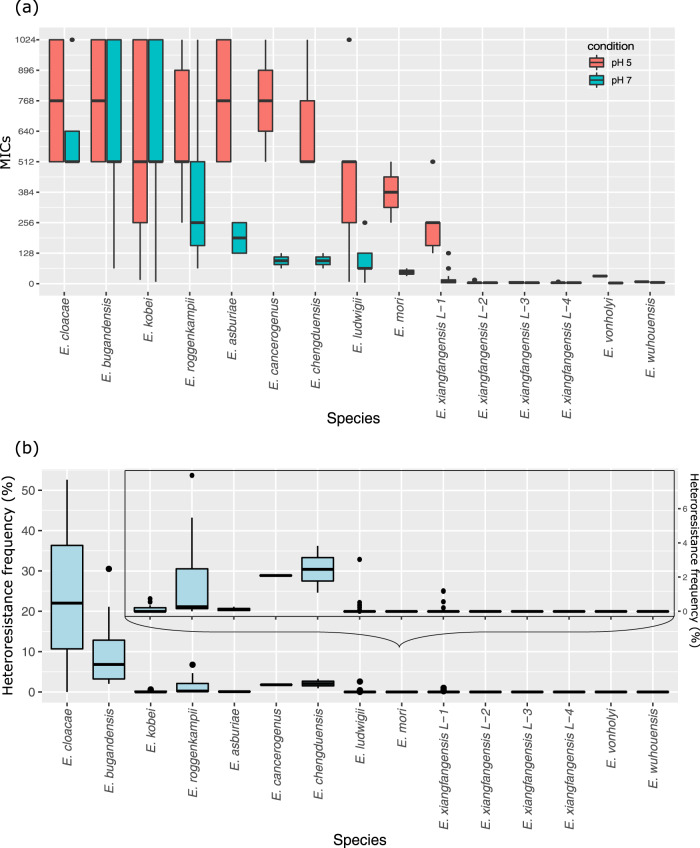
Species within the Enterobacter cloacae complex (ECC) include globally important nosocomial pathogens. A three-year study of ECC in Germany identified Enterobacter xiangfangensis as the most common species (65.5%) detected, a result replicated by examining a global pool of 3246 isolates. Antibiotic resistance profiling revealed widespread resistance and heteroresistance to the antibiotic colistin and detected the mobile colistin resistance (mcr)-9 gene in 19.2% of all isolates. We show that resistance and heteroresistance properties depend on the chromosomal arnBCADTEF gene cassette whose products catalyze transfer of L-Ara4N to lipid A. Using comparative genomics, mutational analysis, and quantitative lipid A profiling we demonstrate that intrinsic lipid A modification levels are genospecies-dependent and governed by allelic variations in phoPQ and mgrB, that encode a two-component sensor-activator system and specific inhibitor peptide. By generating phoPQ chimeras and combining them with mgrB alleles, we show that interactions at the pH-sensing interface of the sensory histidine kinase phoQ dictate arnBCADTEF expression levels. To minimize therapeutic failures, we developed an assay that accurately detects colistin resistance levels for any ECC isolate.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Diekema DJ, Pfaller MA, Jones RN, Beach M. Survey of bloodstream infections due to Gram-negative bacilli: frequency of occurrence and antimicrobial susceptibility of isolates collected in the United States, Canada, and Latin America for the SENTRY antimicrobial surveillance program, 1997. Clin. Infect. Dis. 1999;29:595–607. doi: 10.1086/598640. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
