

A Germline-Targeting Chimpanzee SIV Envelope Glycoprotein Elicits a New Class of V2-Apex Directed Cross-Neutralizing Antibodies
- PMID: 36629414
- PMCID: PMC9973348
- DOI: 10.1128/mbio.03370-22
A Germline-Targeting Chimpanzee SIV Envelope Glycoprotein Elicits a New Class of V2-Apex Directed Cross-Neutralizing Antibodies
Abstract
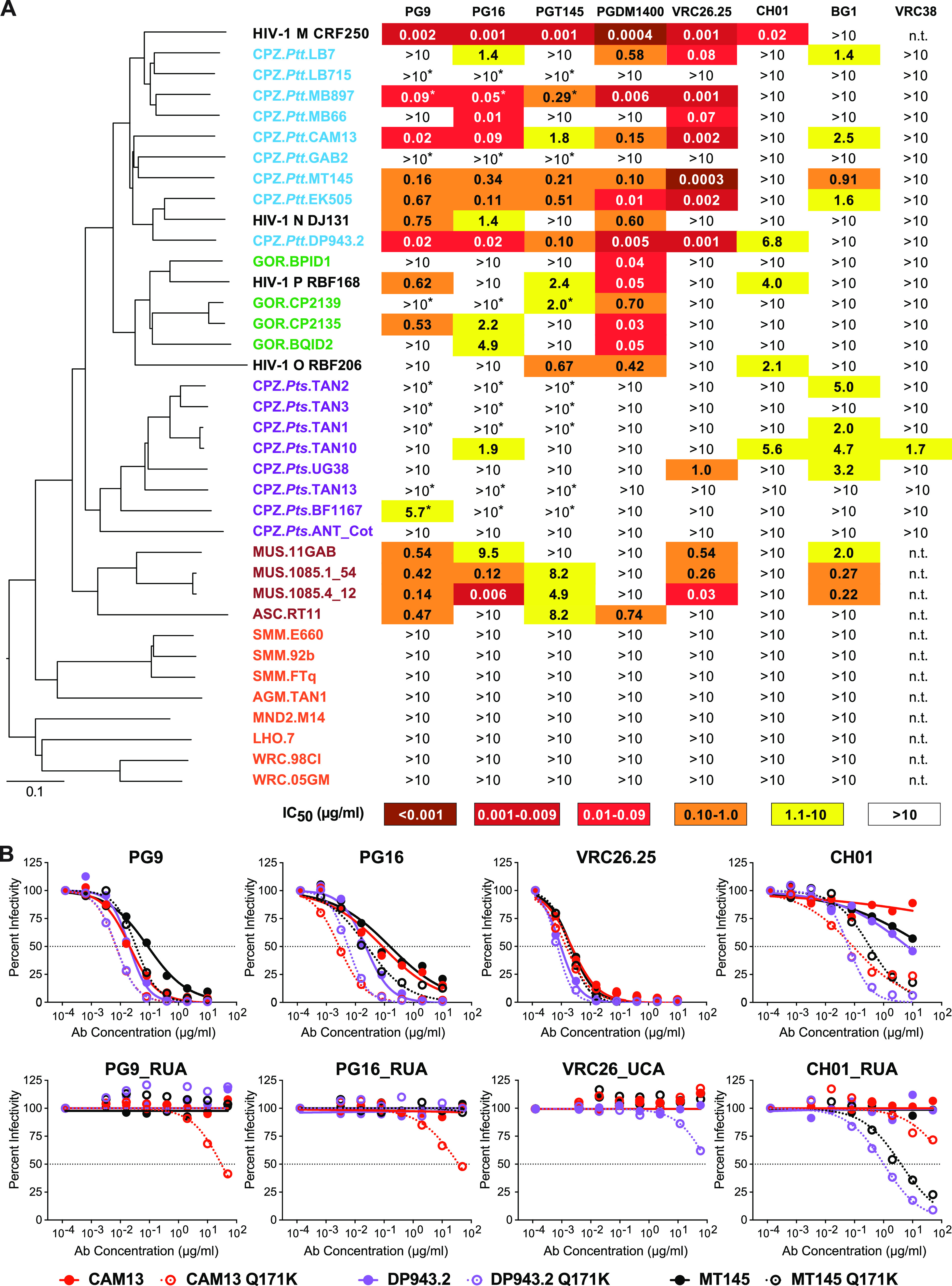
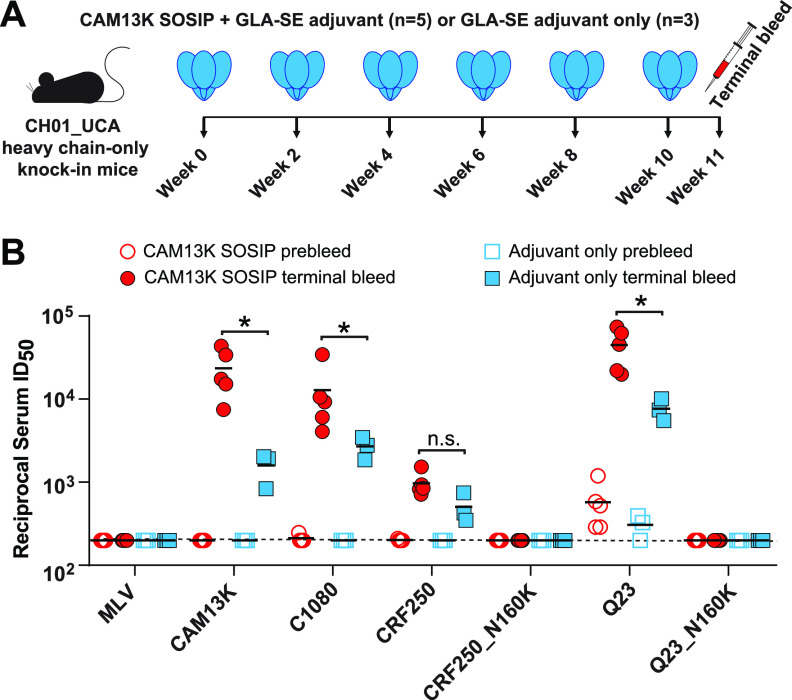
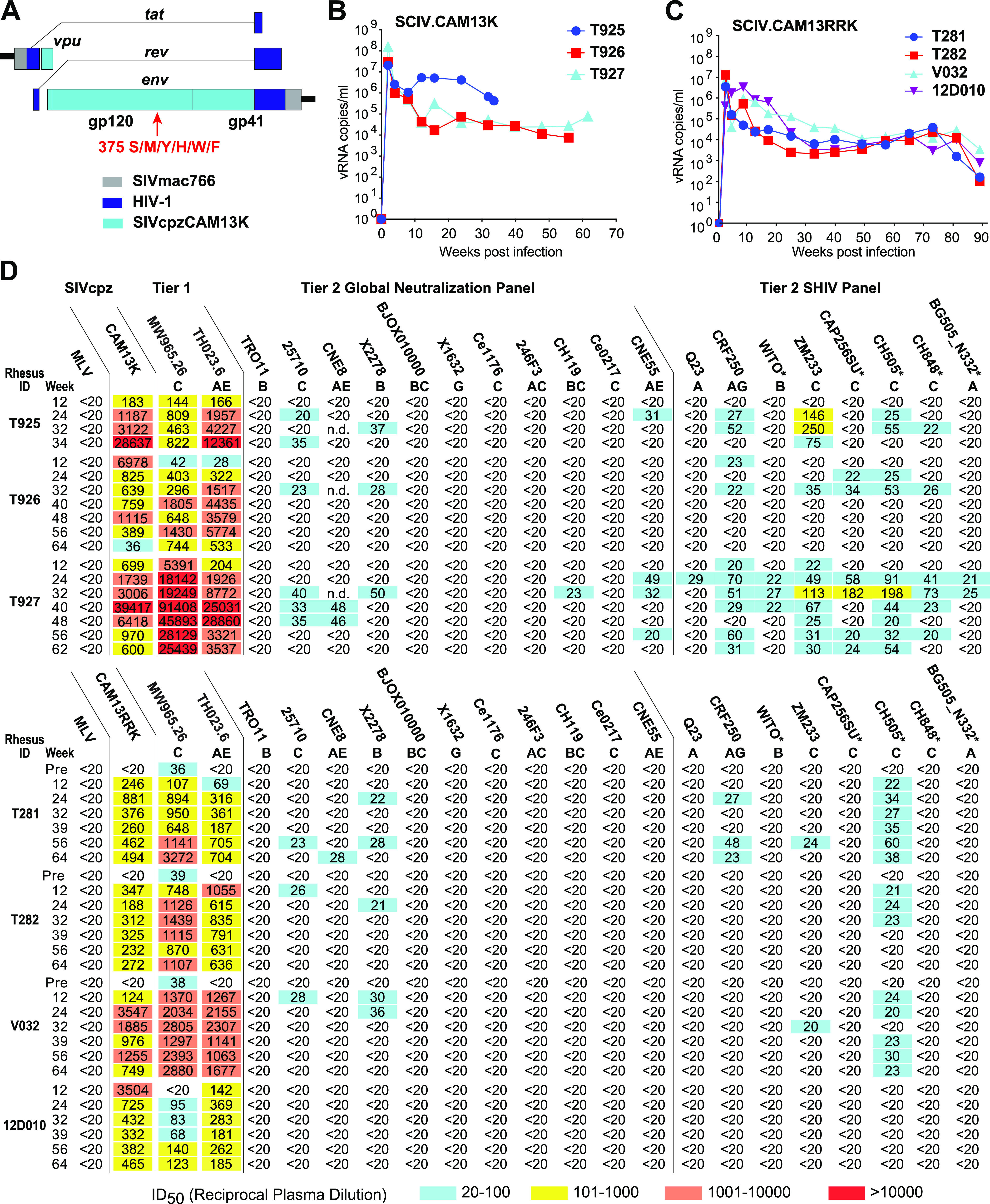
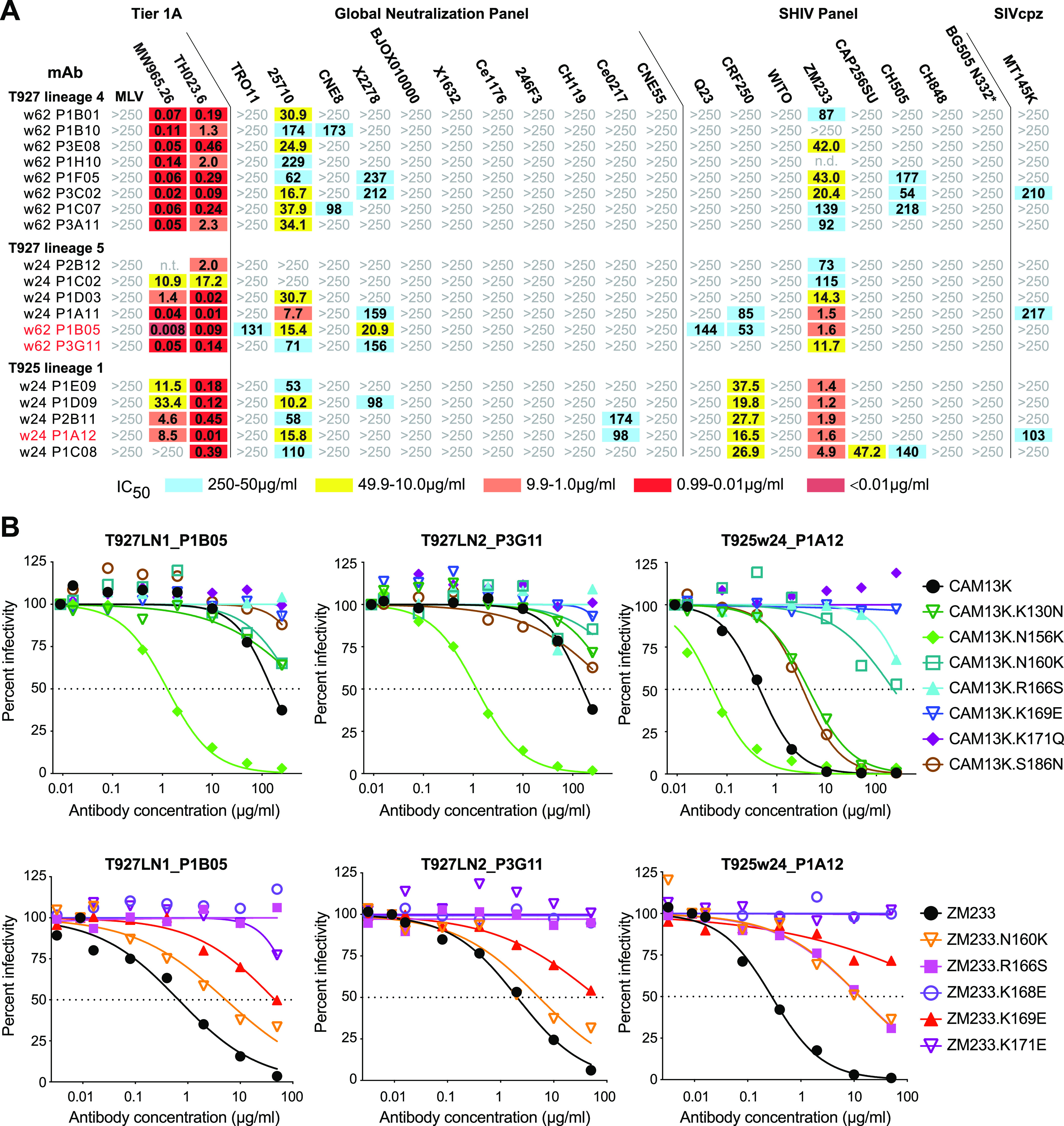
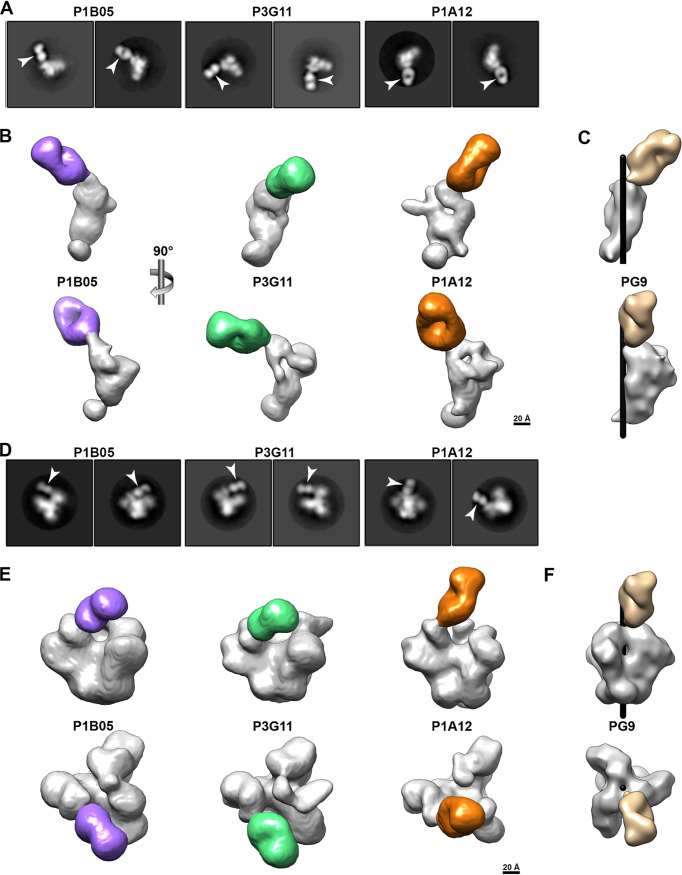
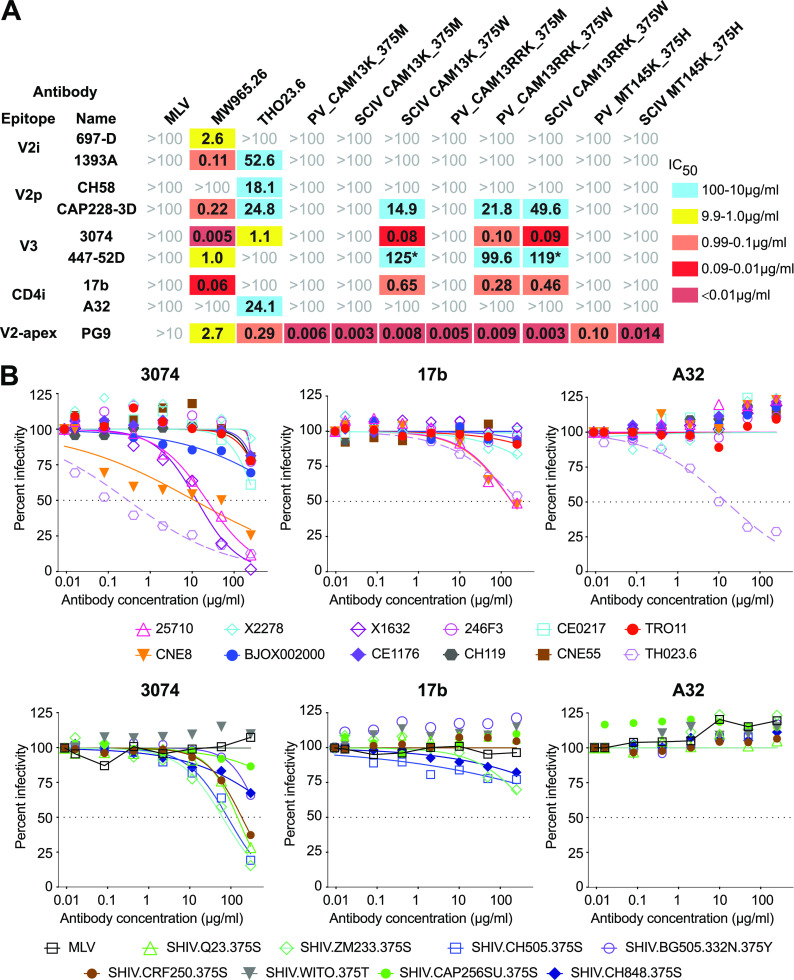
HIV-1 and its SIV precursors share a broadly neutralizing antibody (bNAb) epitope in variable loop 2 (V2) at the envelope glycoprotein (Env) trimer apex. Here, we tested the immunogenicity of germ line-targeting versions of a chimpanzee SIV (SIVcpz) Env in human V2-apex bNAb heavy-chain precursor-expressing knock-in mice and as chimeric simian-chimpanzee immunodeficiency viruses (SCIVs) in rhesus macaques (RMs). Trimer immunization of knock-in mice induced V2-directed NAbs, indicating activation of V2-apex bNAb precursor-expressing mouse B cells. SCIV infection of RMs elicited high-titer viremia, potent autologous tier 2 neutralizing antibodies, and rapid sequence escape in the canonical V2-apex epitope. Six of seven animals also developed low-titer heterologous plasma breadth that mapped to the V2-apex. Antibody cloning from two of these animals identified multiple expanded lineages with long heavy chain third complementarity determining regions that cross-neutralized as many as 7 of 19 primary HIV-1 strains, but with low potency. Negative stain electron microscopy (NSEM) of members of the two most cross-reactive lineages confirmed V2 targeting but identified an angle of approach distinct from prototypical V2-apex bNAbs, with antibody binding either requiring or inducing an occluded-open trimer. Probing with conformation-sensitive, nonneutralizing antibodies revealed that SCIV-expressed, but not wild-type SIVcpz Envs, as well as a subset of primary HIV-1 Envs, preferentially adopted a more open trimeric state. These results reveal the existence of a cryptic V2 epitope that is exposed in occluded-open SIVcpz and HIV-1 Env trimers and elicits cross-neutralizing responses of limited breadth and potency. IMPORTANCE An effective HIV-1 vaccination strategy will need to stimulate rare precursor B cells of multiple bNAb lineages and affinity mature them along desired pathways. Here, we searched for V2-apex germ line-targeting Envs among a large set of diverse primate lentiviruses and identified minimally modified versions of one chimpanzee SIV Env that bound several human V2-apex bNAb precursors and stimulated one of these in a V2-apex bNAb precursor-expressing knock-in mouse. We also generated chimeric simian-chimpanzee immunodeficiency viruses and showed that they elicit low-titer V2-directed heterologous plasma breadth in six of seven infected rhesus macaques. Characterization of this antibody response identified a new class of weakly cross-reactive neutralizing antibodies that target the V2-apex, but only in occluded-open Env trimers. The existence of this cryptic epitope, which in some Env backgrounds is immunodominant, needs to be considered in immunogen design.
Keywords: SCIV; V2-apex; broadly neutralizing antibodies; chimpanzee; germline-targeting; human immunodeficiency virus; immunofocusing; neutralizing antibodies; occluded-open trimer; vaccines.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Kong R, Duan H, Sheng Z, Xu K, Acharya P, Chen X, Cheng C, Dingens AS, Gorman J, Sastry M, Shen C-H, Zhang B, Zhou T, Chuang G-Y, Chao CW, Gu Y, Jafari AJ, Louder MK, O'Dell S, Rowshan AP, Viox EG, Wang Y, Choi CW, Corcoran MM, Corrigan AR, Dandey VP, Eng ET, Geng H, Foulds KE, Guo Y, Kwon YD, Lin B, Liu K, Mason RD, Nason MC, Ohr TY, Ou L, Rawi R, Sarfo EK, Schön A, Todd JP, Wang S, Wei H, Wu W, Mullikin JC, Bailer RT, Doria-Rose NA, Karlsson Hedestam GB, Scorpio DG, Overbaugh J, NISC Comparative Sequencing Program , et al. 2019. Antibody lineages with vaccine-induced antigen-binding hotspots develop broad HIV neutralization. Cell 178:567–584.e19. doi: 10.1016/j.cell.2019.06.030. - DOI - PMC - PubMed
-
- Gray ES, Madiga MC, Hermanus T, Moore PL, Wibmer CK, Tumba NL, Werner L, Mlisana K, Sibeko S, Williamson C, Abdool Karim SS, Morris L, Team CS, CAPRISA002 Study Team . 2011. The neutralization breadth of HIV-1 develops incrementally over four years and is associated with CD4+ T cell decline and high viral load during acute infection. J Virol 85:4828–4840. doi: 10.1128/JVI.00198-11. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
- T32 AI007392/AI/NIAID NIH HHS/United States
- UM1 AI144462/AI/NIAID NIH HHS/United States
- R01 AI167716/AI/NIAID NIH HHS/United States
- R61 AI161818/AI/NIAID NIH HHS/United States
- R01 AI160607/AI/NIAID NIH HHS/United States
- P01 AI131251/AI/NIAID NIH HHS/United States
- U42 OD021458/OD/NIH HHS/United States
- T32 AI007632/AI/NIAID NIH HHS/United States
- R01 AI141338/AI/NIAID NIH HHS/United States
- R01 AI087202/AI/NIAID NIH HHS/United States
- R37 AI150590/AI/NIAID NIH HHS/United States
- R01 AI165080/AI/NIAID NIH HHS/United States
- R01 AI050529/AI/NIAID NIH HHS/United States
- UM1 AI144371/AI/NIAID NIH HHS/United States
- P40 OD012217/OD/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
