

Neuronal membrane proteasomes regulate neuronal circuit activity in vivo and are required for learning-induced behavioral plasticity
- PMID: 36630455
- PMCID: PMC9934054
- DOI: 10.1073/pnas.2216537120
Neuronal membrane proteasomes regulate neuronal circuit activity in vivo and are required for learning-induced behavioral plasticity
Abstract
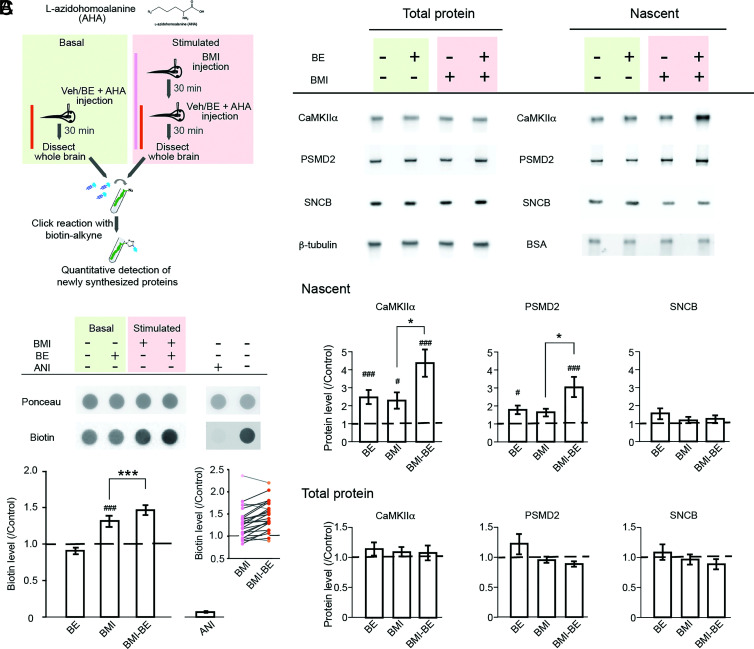
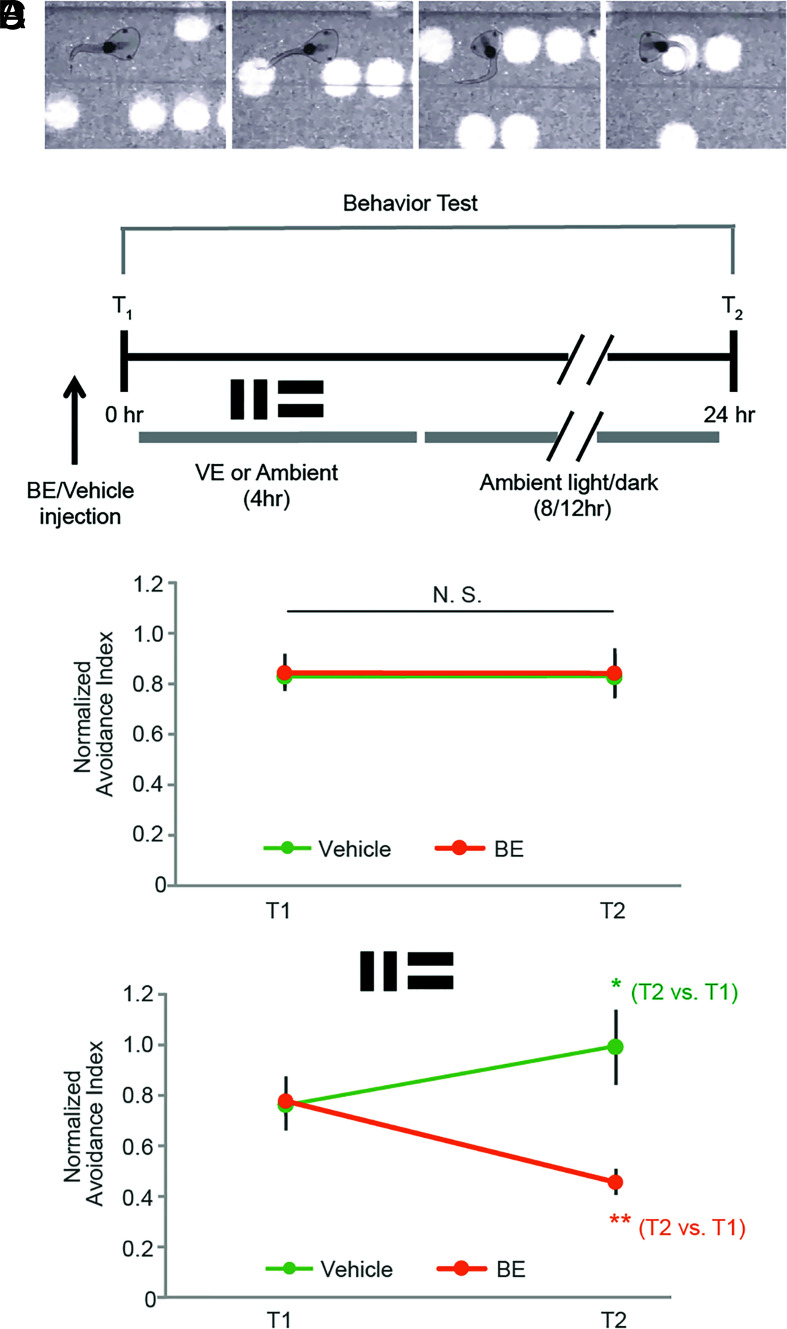
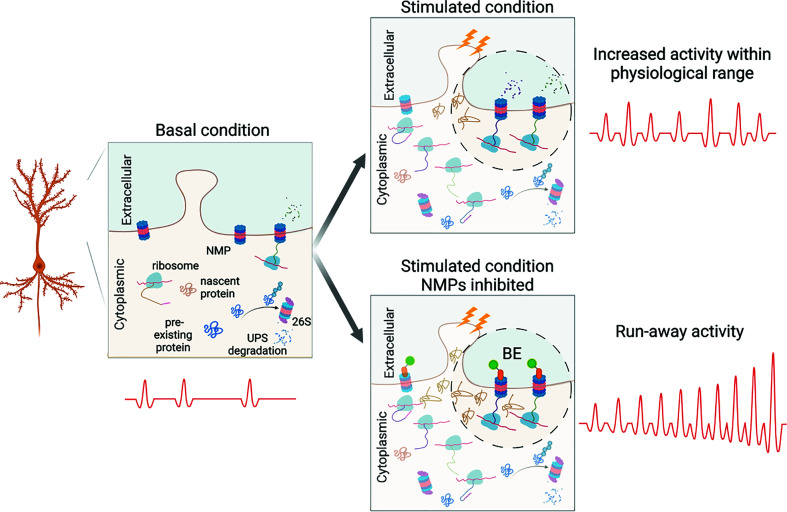
Protein degradation is critical for brain function through processes that remain incompletely understood. Here, we investigated the in vivo function of the 20S neuronal membrane proteasome (NMP) in the brain of Xenopus laevis tadpoles. With biochemistry, immunohistochemistry, and electron microscopy, we demonstrated that NMPs are conserved in the tadpole brain and preferentially degrade neuronal activity-induced newly synthesized proteins in vivo. Using in vivo calcium imaging in the optic tectum, we showed that acute NMP inhibition rapidly increased spontaneous neuronal activity, resulting in hypersynchronization across tectal neurons. At the circuit level, inhibiting NMPs abolished learning-dependent improvement in visuomotor behavior in live animals and caused a significant deterioration in basal behavioral performance following visual training with enhanced visual experience. Our data provide in vivo characterization of NMP functions in the vertebrate nervous system and suggest that NMP-mediated degradation of activity-induced nascent proteins may serve as a homeostatic modulatory mechanism in neurons that is critical for regulating neuronal activity and experience-dependent circuit plasticity.
Keywords: BONCAT; activity-induced nascent proteins; neuron; proteasome; spontaneous activity.
Conflict of interest statement
The authors declare no competing interest.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
