

Activated regulatory T-cells promote duodenal bacterial translocation into necrotic areas in severe acute pancreatitis
- PMID: 36631247
- PMCID: PMC10314084
- DOI: 10.1136/gutjnl-2022-327448
Activated regulatory T-cells promote duodenal bacterial translocation into necrotic areas in severe acute pancreatitis
Abstract
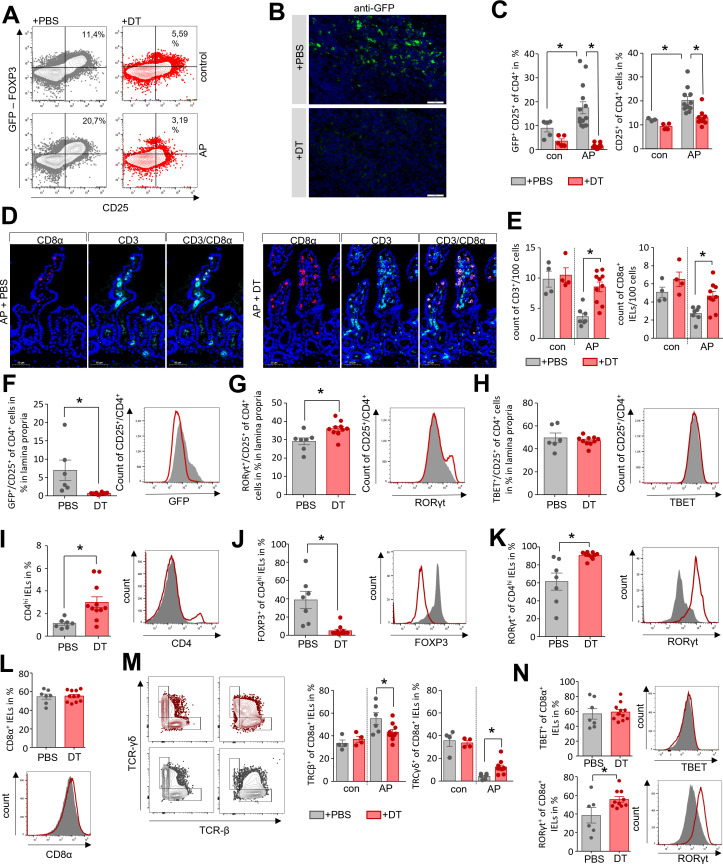
Objective: In acute pancreatitis (AP), bacterial translocation and subsequent infection of pancreatic necrosis are the main risk factors for severe disease and late death. Understanding how immunological host defence mechanisms fail to protect the intestinal barrier is of great importance in reducing the mortality risk of the disease. Here, we studied the role of the Treg/Th17 balance for maintaining the intestinal barrier function in a mouse model of severe AP.
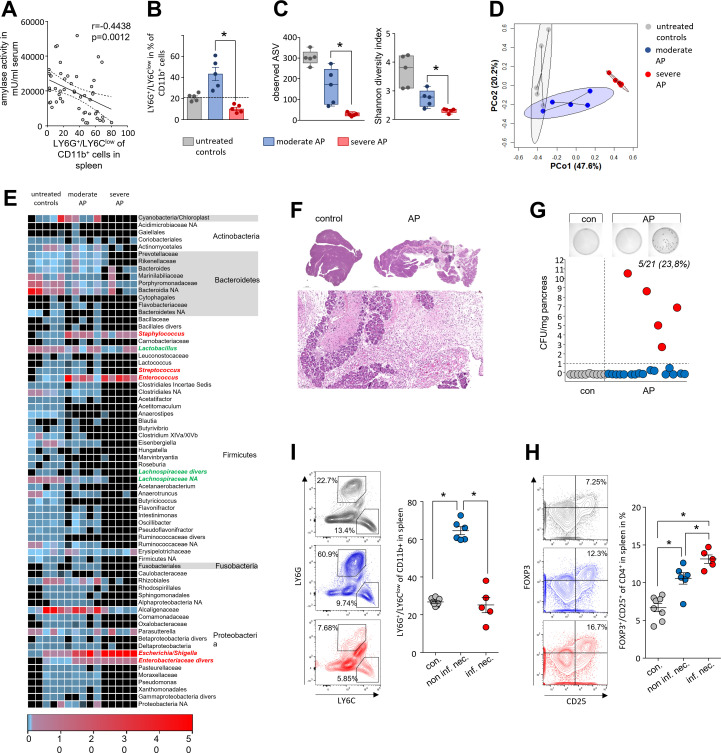
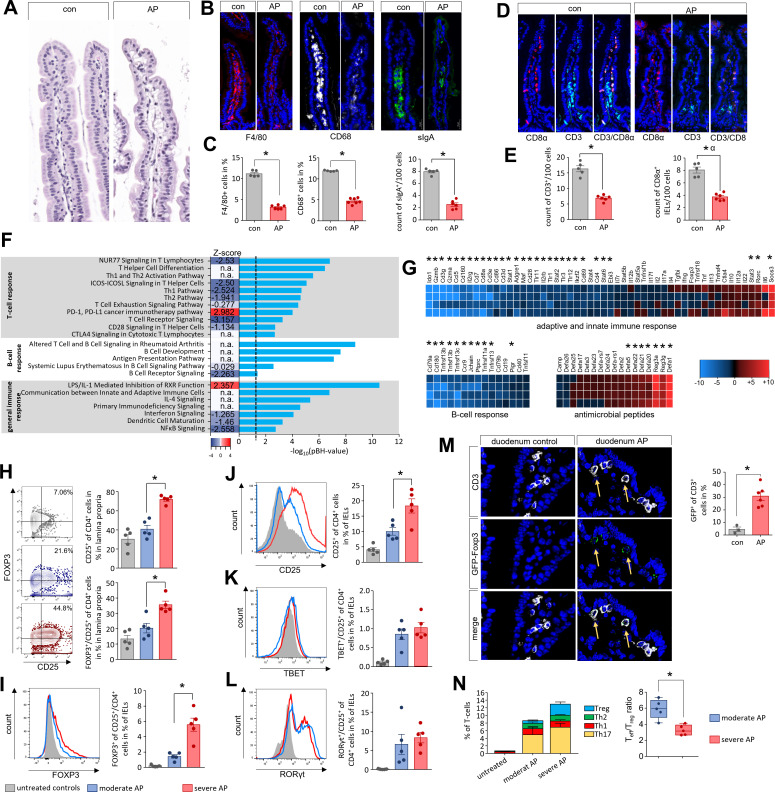
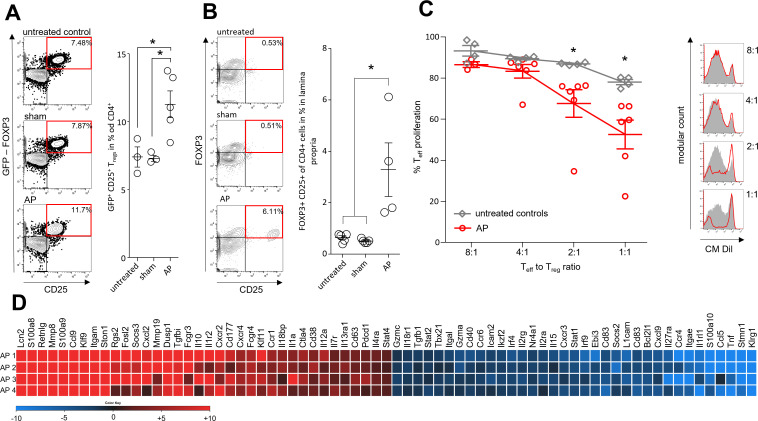
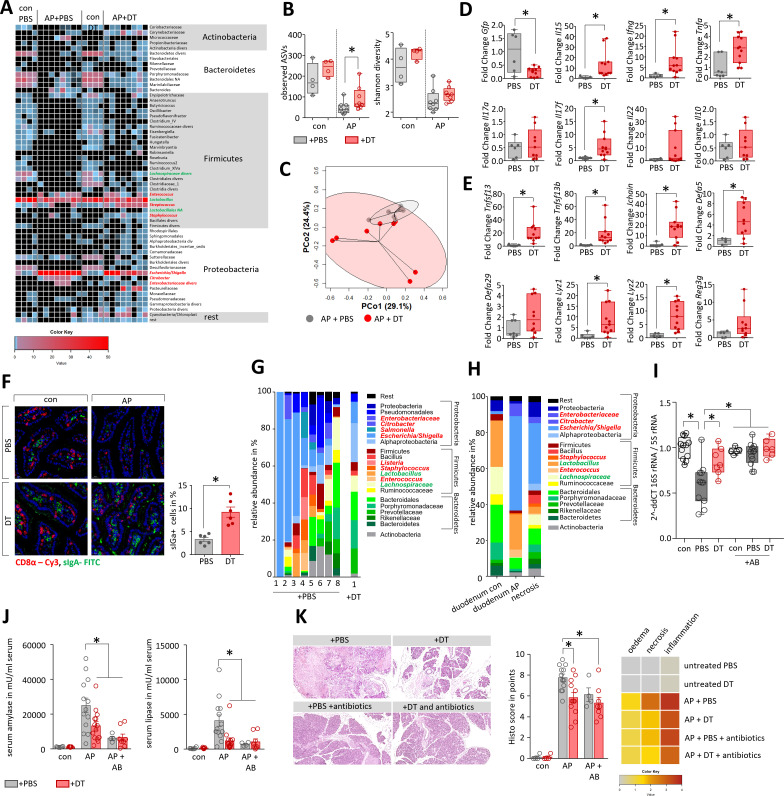
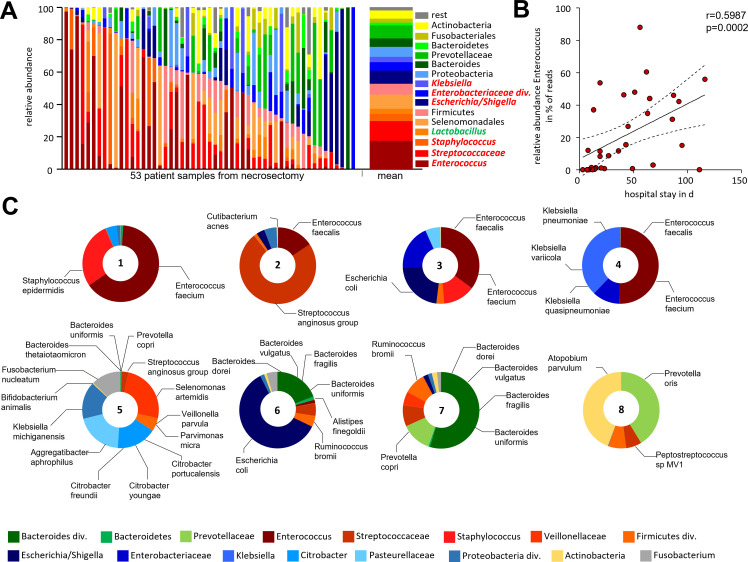
Design: AP was induced by partial duct ligation in C57Bl/6 or DEREG mice, in which regulatory T-cells (Treg) were depleted by intraperitoneal injection of diphtheria toxin. By flow cytometry, functional suppression assays and transcriptional profiling we analysed Treg activation and characterised T-cells of the lamina propria as well as intraepithelial lymphocytes (IELs) regarding their activation and differentiation. Microbiota composition was examined in intestinal samples as well as in murine and human pancreatic necrosis by 16S rRNA gene sequencing.
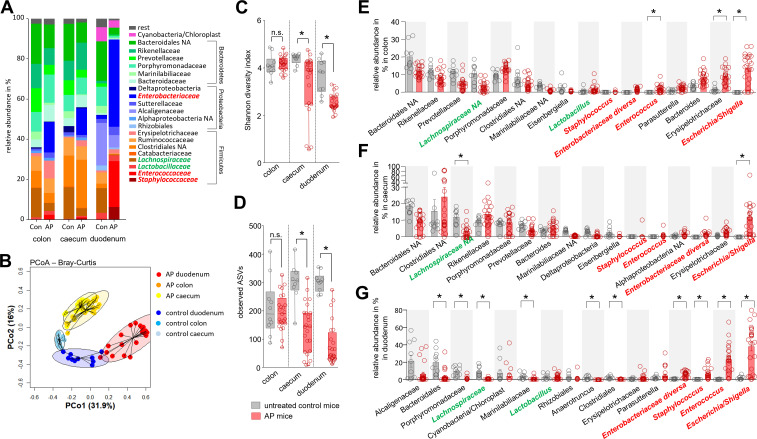
Results: The prophylactic Treg-depletion enhanced the proinflammatory response in an experimental mouse model of AP but stabilised the intestinal immunological barrier function of Th17 cells and CD8+/γδTCR+ IELs. Treg depleted animals developed less bacterial translocation to the pancreas. Duodenal overgrowth of the facultative pathogenic taxa Escherichia/Shigella which associates with severe disease and infected necrosis was diminished in Treg depleted animals.
Conclusion: Tregs play a crucial role in the counterbalance against systemic inflammatory response syndrome. In AP, Treg-activation disturbs the duodenal barrier function and permits translocation of commensal bacteria into pancreatic necrosis. Targeting Tregs in AP may help to ameliorate the disease course.
Keywords: acute pancreatitis; bacterial infection; experimental pancreatitis; immune response.
© Author(s) (or their employer(s)) 2023. Re-use permitted under CC BY-NC. No commercial re-use. See rights and permissions. Published by BMJ.
Conflict of interest statement
Competing interests: None declared.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials