

Single-nuclei transcriptomics enable detection of somatic variants in patient brain tissue
- PMID: 36631516
- PMCID: PMC9834227
- DOI: 10.1038/s41598-023-27700-6
Single-nuclei transcriptomics enable detection of somatic variants in patient brain tissue
Abstract
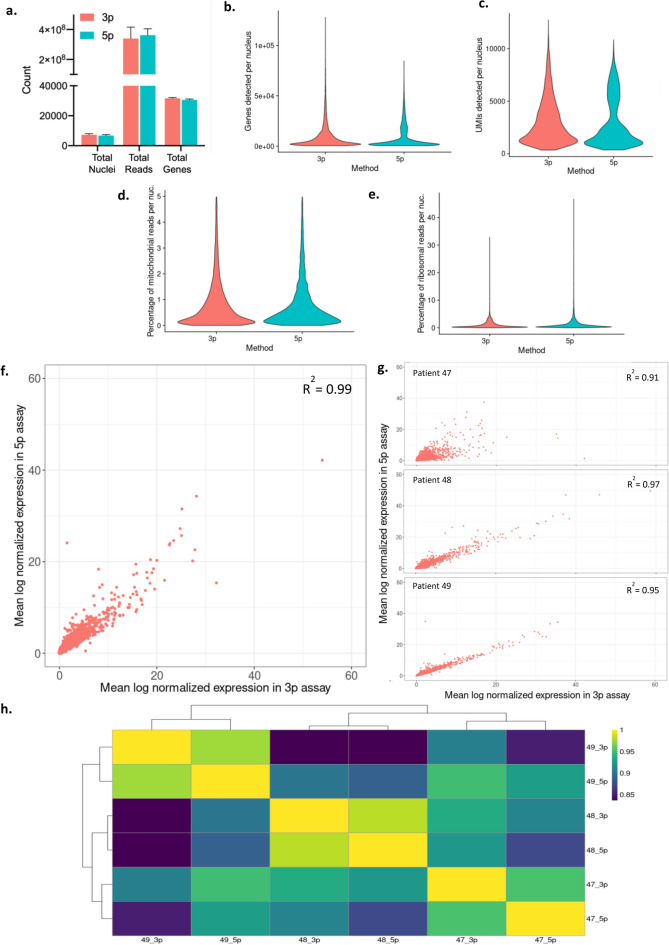
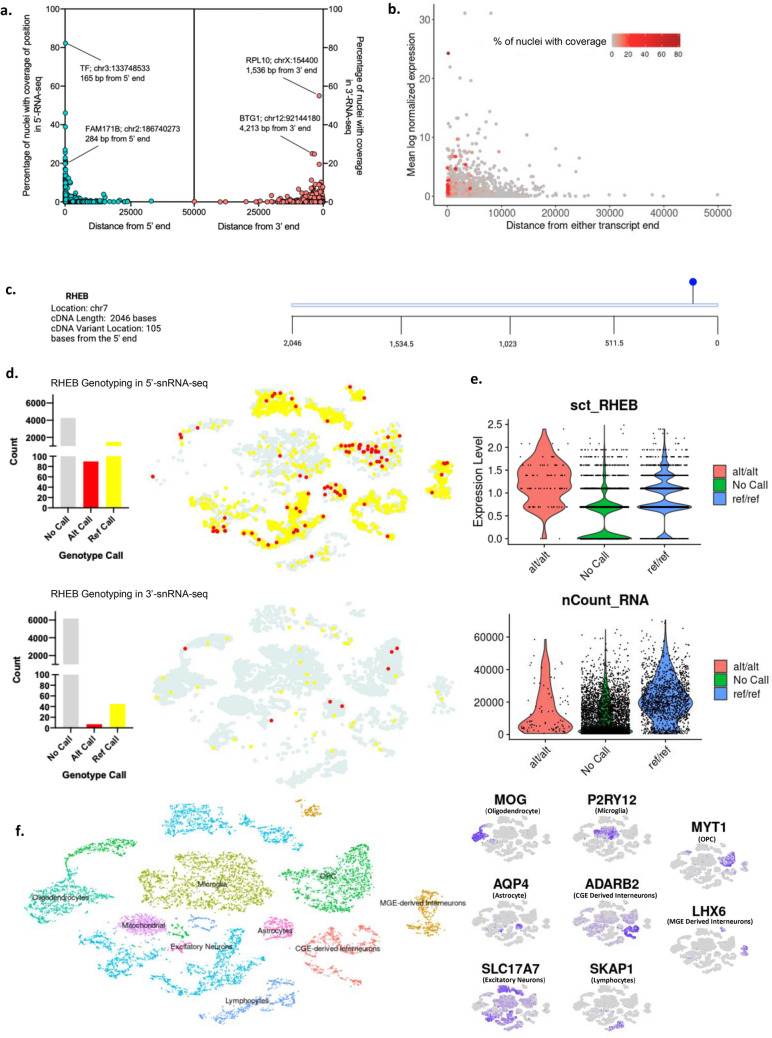
Somatic variants are a major cause of human disease, including neurological disorders like focal epilepsies, but can be challenging to study due to their mosaicism in bulk tissue biopsies. Coupling single-cell genotype and transcriptomic data has potential to provide insight into the role somatic variants play in disease etiology, such as by determining what cell types are affected or how the mutations affect gene expression. Here, we asked whether commonly used single-nucleus 3'- or 5'-RNA-sequencing assays can be used to derive single-nucleus genotype data for a priori known variants that are located near to either end of a transcript. To that end, we compared performance of commercially available single-nuclei 3'- and 5'- gene expression kits using resected brain samples from three pediatric patients with focal epilepsy. We quantified the ability to detect genetic variants in single-nucleus datasets depending on distance from the transcript end. Finally, we demonstrated the ability to identify affected cell types in a patient with a RHEB somatic variant causing an epilepsy-associated cortical malformation. Our results demonstrate that single-nuclei 3' or 5'-RNA-sequencing data can be used to identify known somatic variants in single-nuclei when they are expressed within proximity to a transcript end.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
