

Lateral Septum Somatostatin Neurons are Activated by Diverse Stressors
- PMID: 36631846
- PMCID: PMC9841747
- DOI: 10.5607/en22024
Lateral Septum Somatostatin Neurons are Activated by Diverse Stressors
Abstract
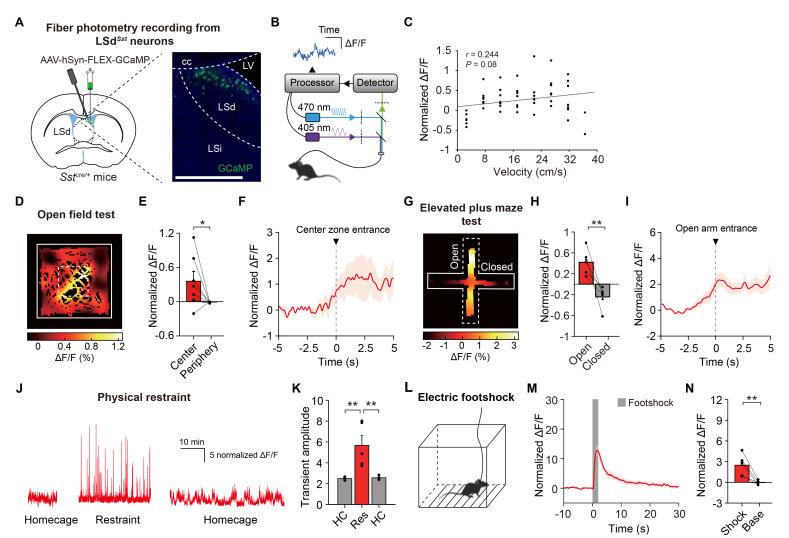
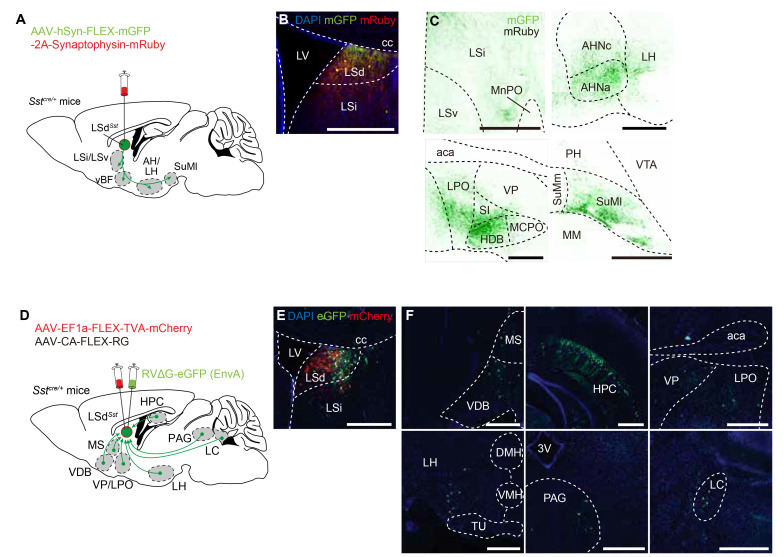
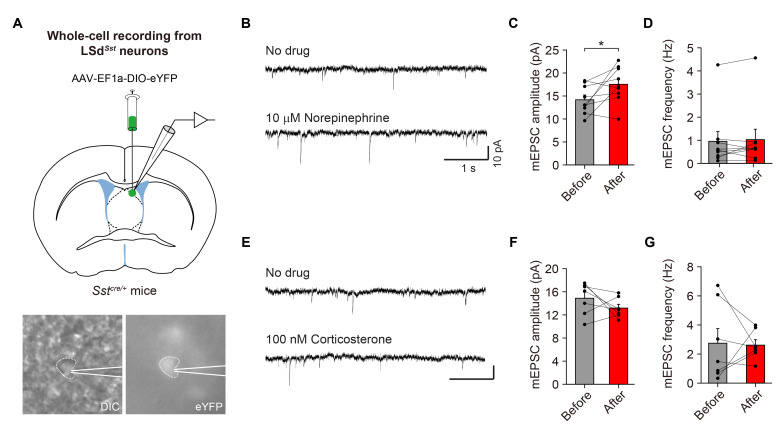
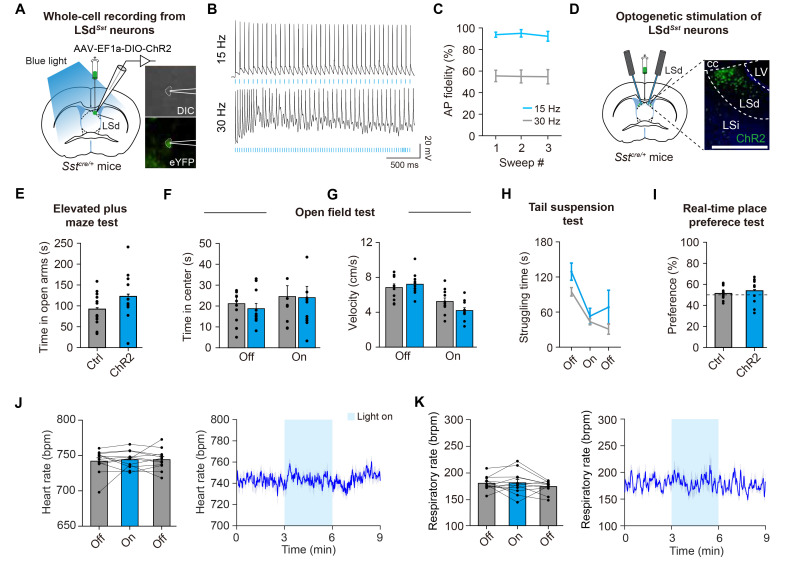
The lateral septum (LS) is a forebrain structure that has been implicated in a wide range of behavioral and physiological responses to stress. However, the specific populations of neurons in the LS that mediate stress responses remain incompletely understood. Here, we show that neurons in the dorsal lateral septum (LSd) that express the somatostatin gene (hereafter, LSdSst neurons) are activated by diverse stressors. Retrograde tracing from LSdSst neurons revealed that these neurons are directly innervated by neurons in the locus coeruleus (LC), the primary source of norepinephrine well-known to mediate diverse stress-related functions in the brain. Consistently, we found that norepinephrine increased excitatory synaptic transmission onto LSdSst neurons, suggesting the functional connectivity between LSdSst neurons and LC noradrenergic neurons. However, optogenetic stimulation of LSdSst neurons did not affect stress-related behaviors or autonomic functions, likely owing to the functional heterogeneity within this population. Together, our findings show that LSdSst neurons are activated by diverse stressors and suggest that norepinephrine released from the LC may modulate the activity of LSdSst neurons under stressful circumstances.
Keywords: Lateral septum; Locus coeruleus; Norepinephrine; Somatostatin; Stress.
Figures
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
