

Nucleophagy contributes to genome stability through degradation of type II topoisomerases A and B and nucleolar components
- PMID: 36633090
- PMCID: PMC10112964
- DOI: 10.1242/jcs.260563
Nucleophagy contributes to genome stability through degradation of type II topoisomerases A and B and nucleolar components
Abstract
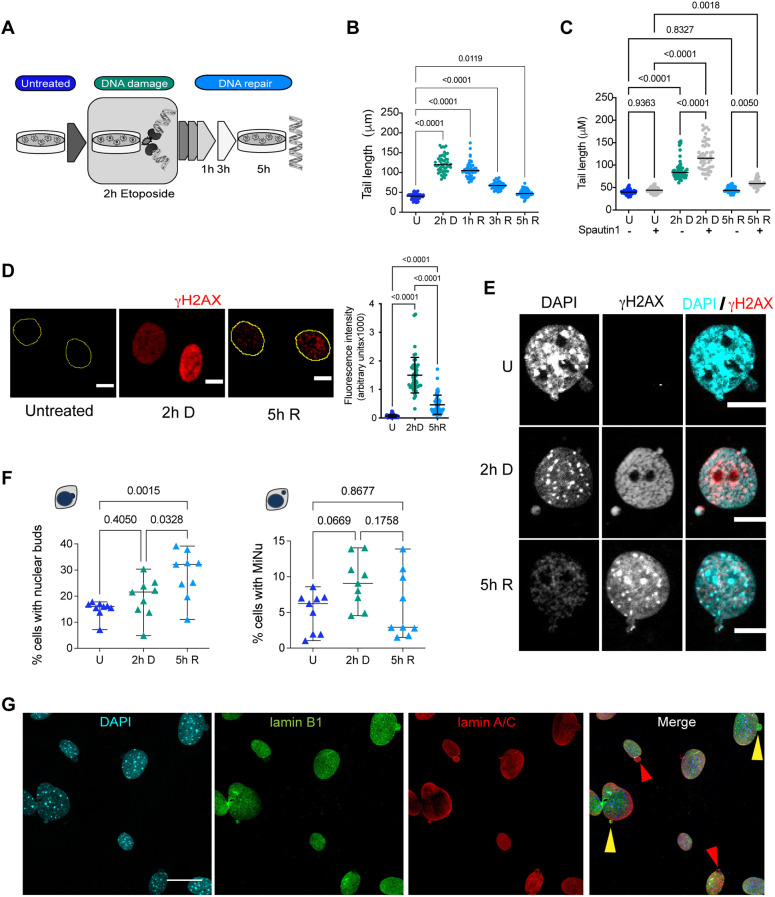
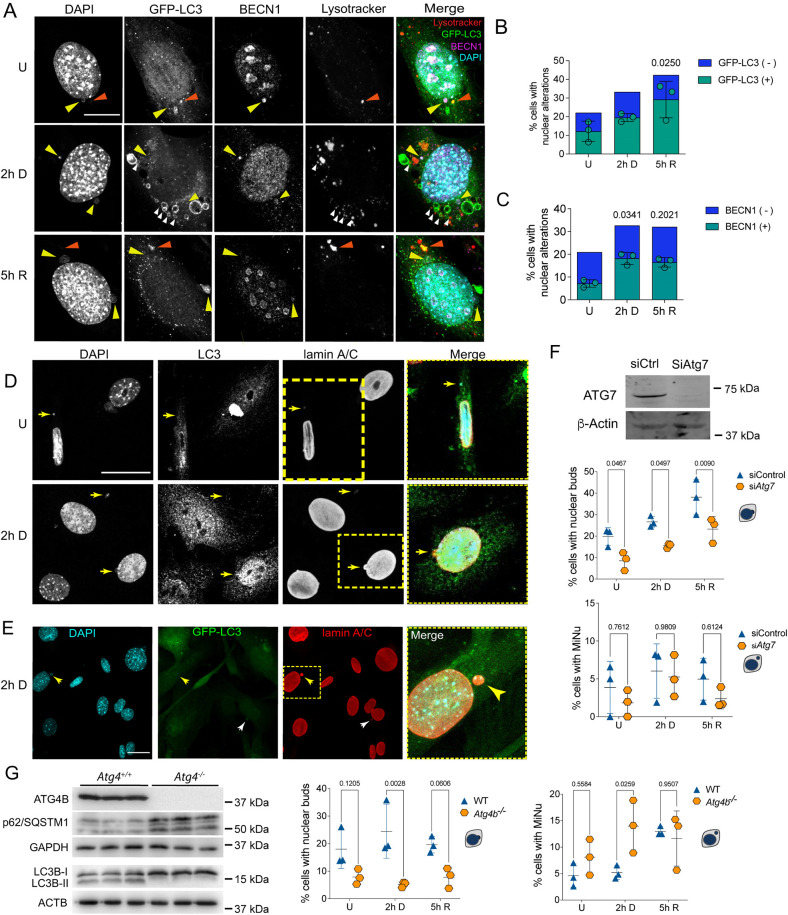
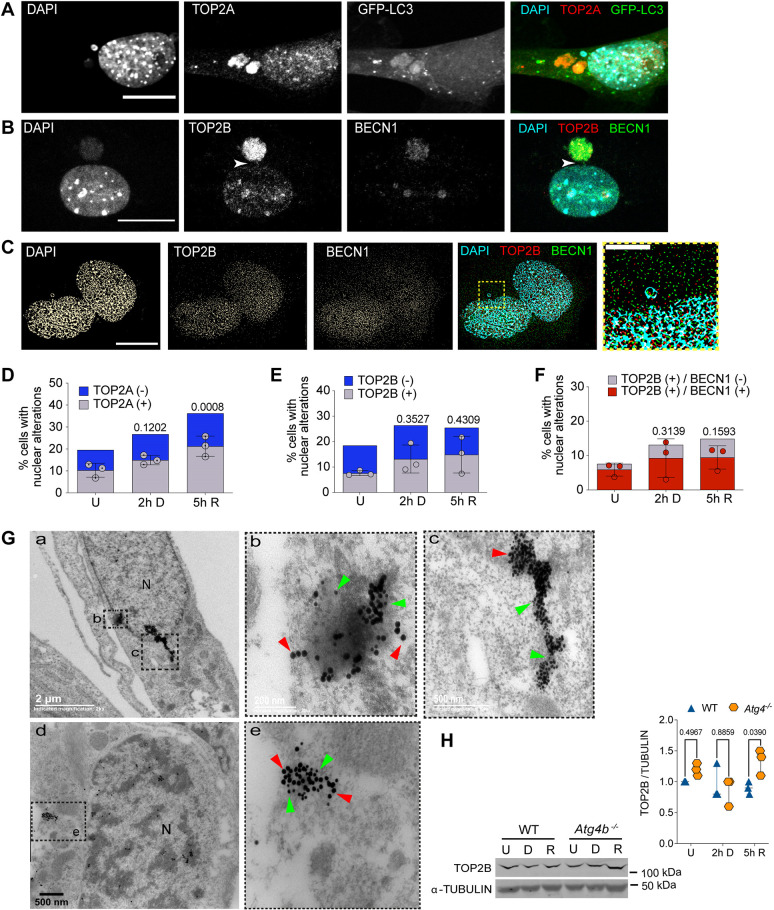
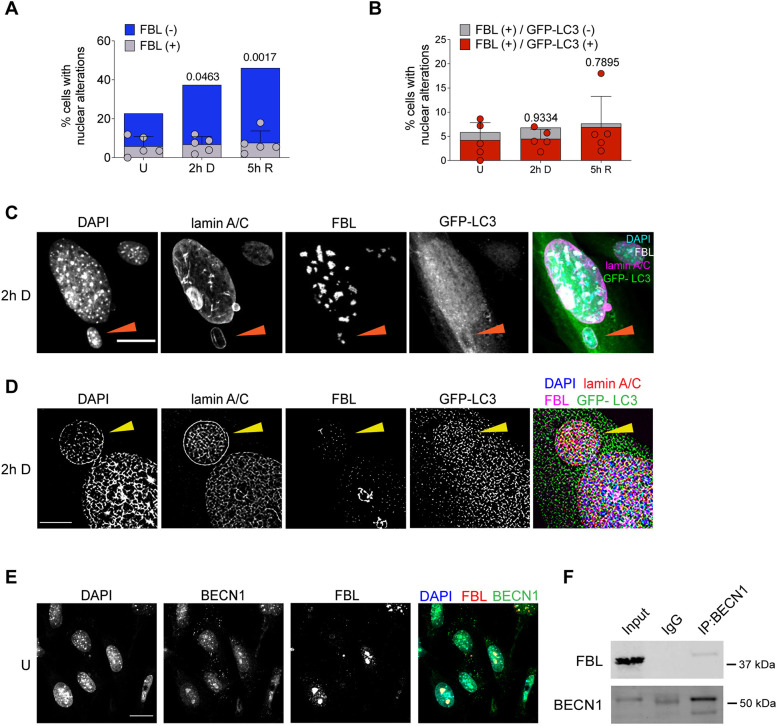
The nuclear architecture of mammalian cells can be altered as a consequence of anomalous accumulation of nuclear proteins or genomic alterations. Most of the knowledge about nuclear dynamics comes from studies on cancerous cells. How normal healthy cells maintain genome stability, avoiding accumulation of nuclear damaged material, is less understood. Here, we describe that primary mouse embryonic fibroblasts develop a basal level of nuclear buds and micronuclei, which increase after etoposide-induced DNA double-stranded breaks. Both basal and induced nuclear buds and micronuclei colocalize with the autophagic proteins BECN1 and LC3B (also known as MAP1LC3B) and with acidic vesicles, suggesting their clearance by nucleophagy. Some of the nuclear alterations also contain autophagic proteins and type II DNA topoisomerases (TOP2A and TOP2B), or the nucleolar protein fibrillarin, implying they are also targets of nucleophagy. We propose that basal nucleophagy contributes to genome and nuclear stability, as well as in response to DNA damage.
Keywords: Autophagy; DNA damage; Mammalian nucleophagy; Micronuclei; Nucleolus.
© 2023. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
Similar articles
-
Nuclear accumulation of UBC9 contributes to SUMOylation of lamin A/C and nucleophagy in response to DNA damage.J Exp Clin Cancer Res. 2019 Feb 11;38(1):67. doi: 10.1186/s13046-019-1048-8. J Exp Clin Cancer Res. 2019. PMID: 30744690 Free PMC article.
-
Nucleophagy mediators and mechanisms.Prog Mol Biol Transl Sci. 2020;172:1-14. doi: 10.1016/bs.pmbts.2020.01.003. Epub 2020 Feb 3. Prog Mol Biol Transl Sci. 2020. PMID: 32620238 Review.
-
CLIP and cohibin separate rDNA from nucleolar proteins destined for degradation by nucleophagy.J Cell Biol. 2018 Aug 6;217(8):2675-2690. doi: 10.1083/jcb.201706164. Epub 2018 Jun 29. J Cell Biol. 2018. PMID: 29959231 Free PMC article.
-
Nucleophagy-Implications for Microautophagy and Health.Int J Mol Sci. 2020 Jun 24;21(12):4506. doi: 10.3390/ijms21124506. Int J Mol Sci. 2020. PMID: 32599961 Free PMC article. Review.
-
Carbon and nitrogen depletion-induced nucleophagy and selective autophagic sequestration of a whole nucleus in multinucleate cells of the filamentous fungus Aspergillus oryzae.J Gen Appl Microbiol. 2017 May 12;63(2):139-146. doi: 10.2323/jgam.2016.09.001. Epub 2017 Mar 22. J Gen Appl Microbiol. 2017. PMID: 28331162
Cited by
-
LC3B drives transcription-associated homologous recombination via direct interaction with R-loops.Nucleic Acids Res. 2024 May 22;52(9):5088-5106. doi: 10.1093/nar/gkae156. Nucleic Acids Res. 2024. PMID: 38412240 Free PMC article.
-
Insights from a Computational-Based Approach for Analyzing Autophagy Genes across Human Cancers.Genes (Basel). 2023 Jul 28;14(8):1550. doi: 10.3390/genes14081550. Genes (Basel). 2023. PMID: 37628602 Free PMC article. Review.
-
Origin and Fate of Micronuclei on the Road to Chromoanagenesis.Methods Mol Biol. 2025;2968:361-372. doi: 10.1007/978-1-0716-4750-9_21. Methods Mol Biol. 2025. PMID: 40884655
-
A quantitative ultrastructural timeline of nuclear autophagy reveals a role for dynamin-like protein 1 at the nuclear envelope.Nat Cell Biol. 2025 Mar;27(3):464-476. doi: 10.1038/s41556-025-01612-1. Epub 2025 Feb 7. Nat Cell Biol. 2025. PMID: 39920277 Free PMC article.
-
Types of Cell Death from a Molecular Perspective.Biology (Basel). 2023 Nov 13;12(11):1426. doi: 10.3390/biology12111426. Biology (Basel). 2023. PMID: 37998025 Free PMC article. Review.
References
-
- Alchanati, I., Teicher, C., Cohen, G., Shemesh, V., Barr, H. M., Nakache, P., Ben-Avraham, D., Idelevich, A., Angel, I., Livnah, N.et al. (2009). The E3 ubiquitin-ligase Bmi1/Ring1A controls the proteasomal degradation of Top2alpha cleavage complex - a potentially new drug target. PLoS One 4, e8104. 10.1371/journal.pone.0008104 - DOI - PMC - PubMed
-
- Alt, F. W., Wei, P. C. and Schwer, B. (2017). Recurrently breaking genes in neural progenitors: potential roles of DNA breaks in neuronal function, degeneration and cancer. In Genome Editing in Neurosciences (ed. Jaenisch R., Zhang F. and Gage F.), pp. 63-72. Springer. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
