

Dome1-JAK-STAT signaling between parasite and host integrates vector immunity and development
- PMID: 36634189
- PMCID: PMC10122270
- DOI: 10.1126/science.abl3837
Dome1-JAK-STAT signaling between parasite and host integrates vector immunity and development
Abstract
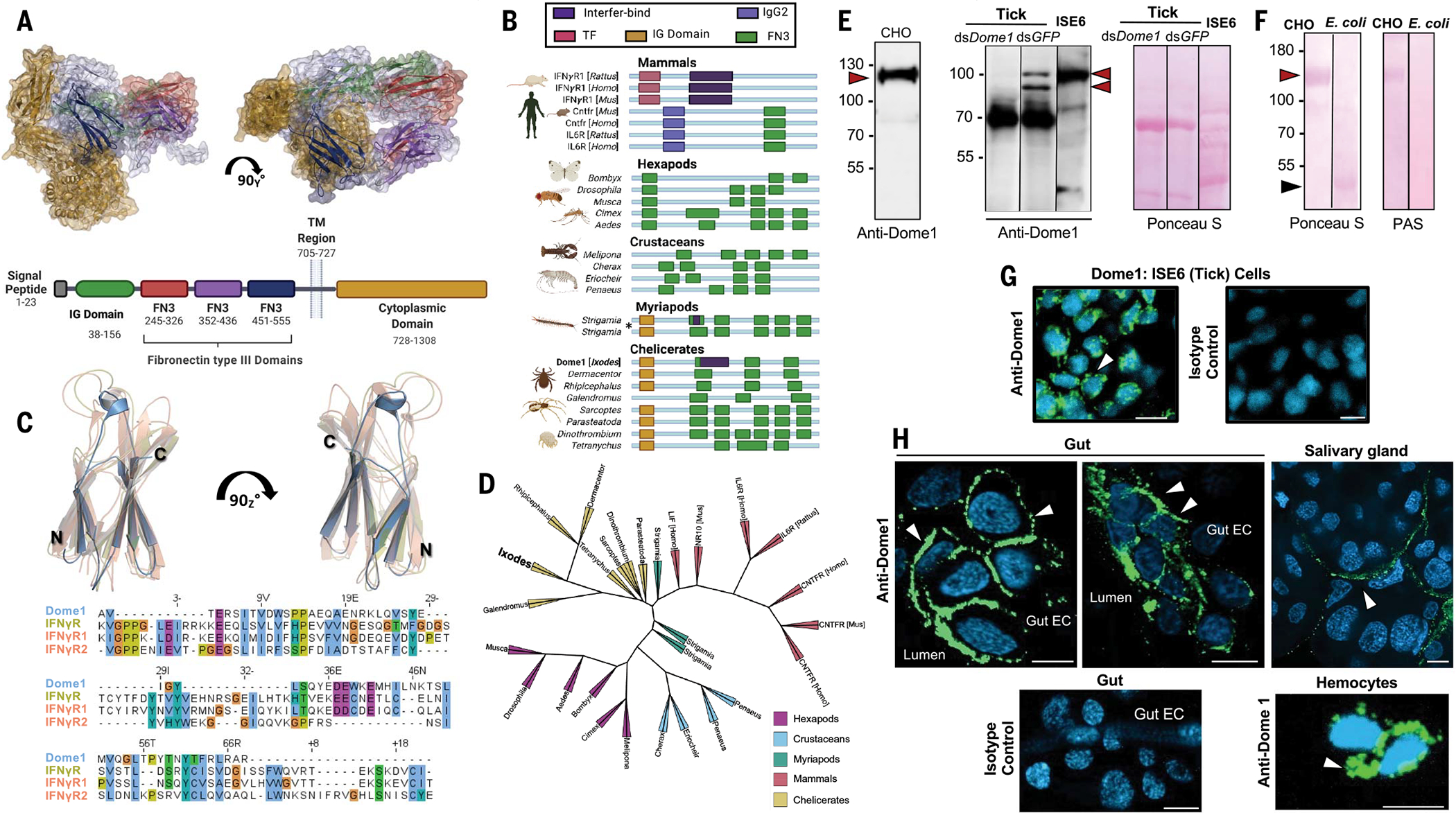
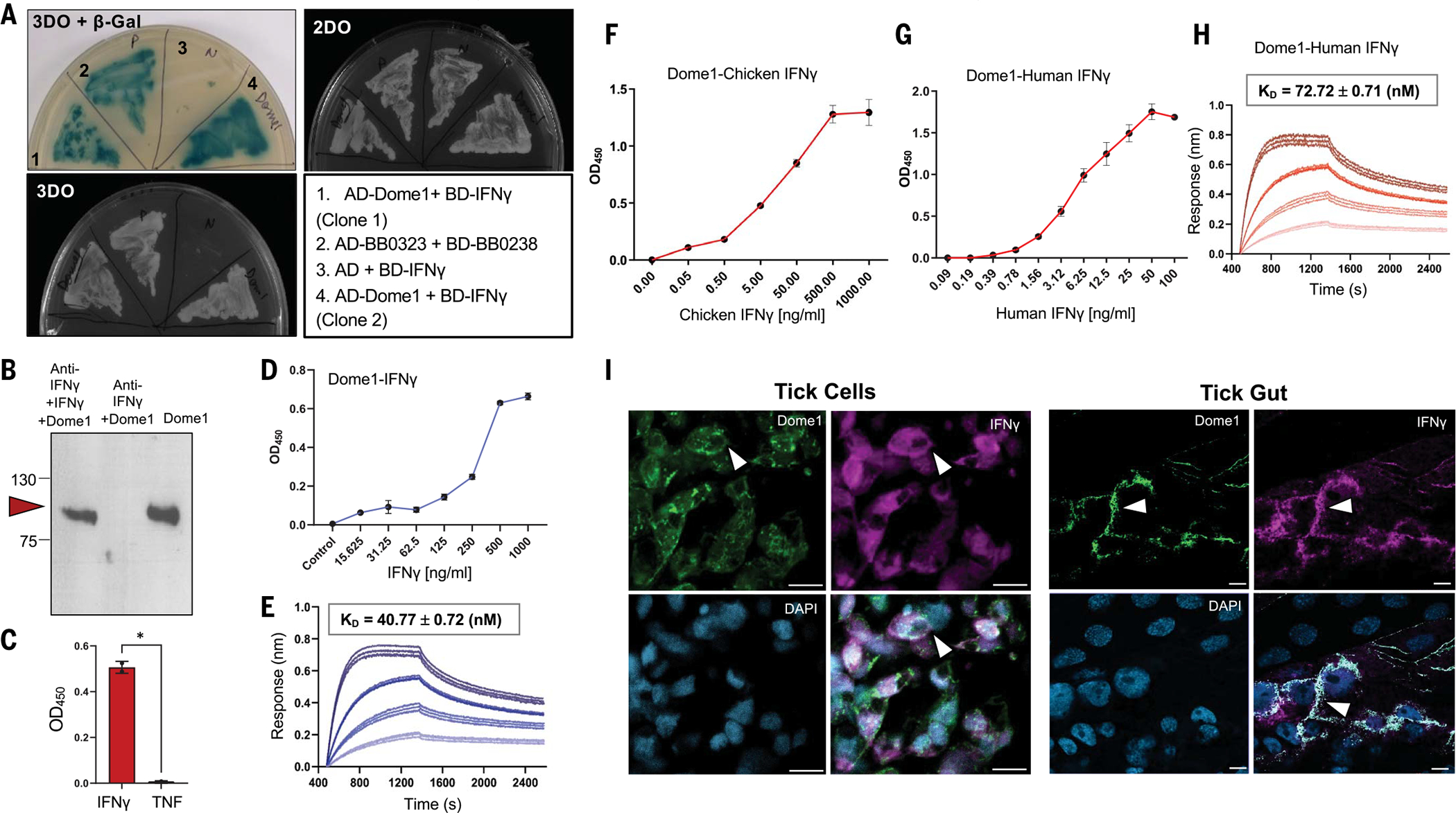
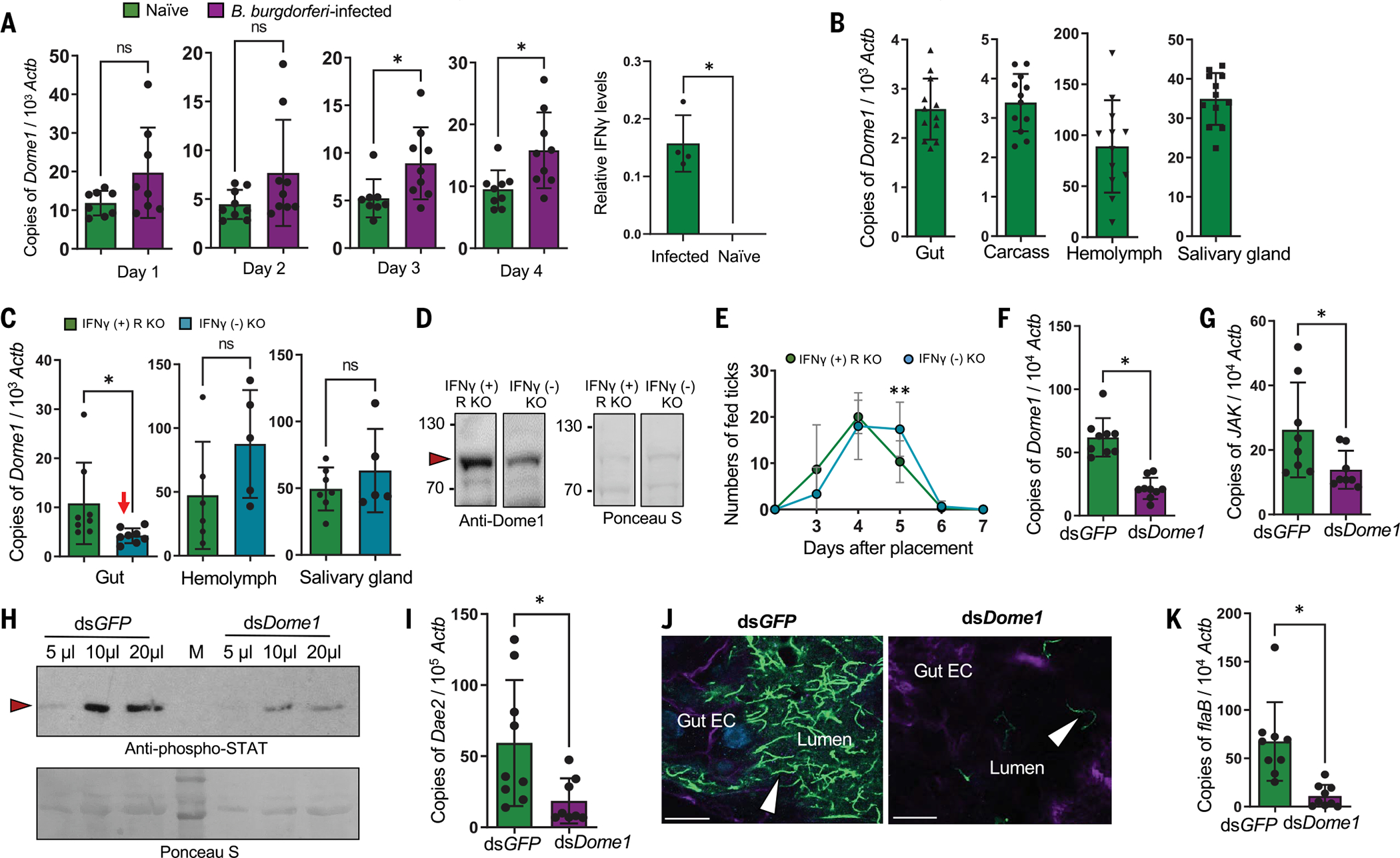
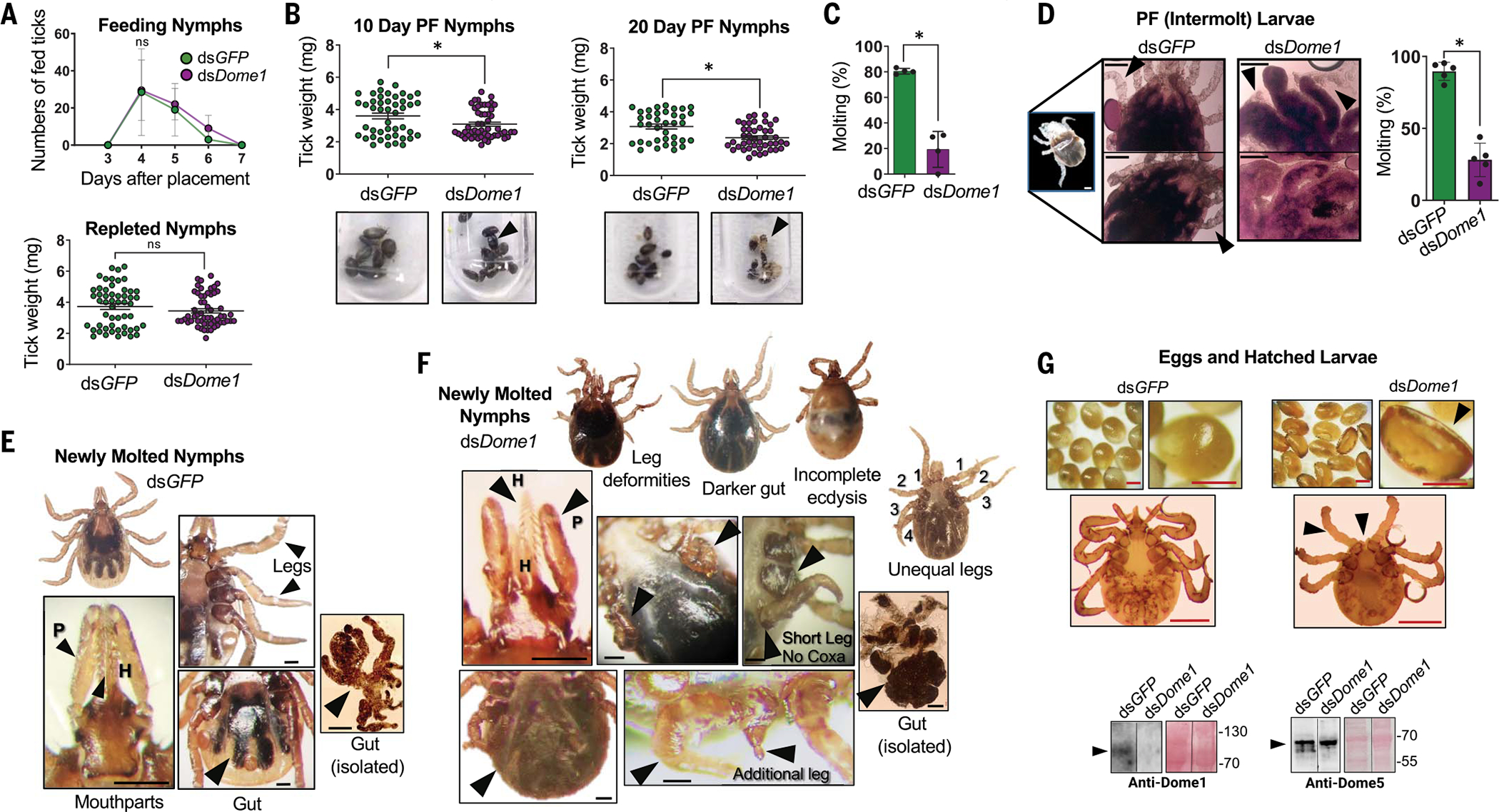
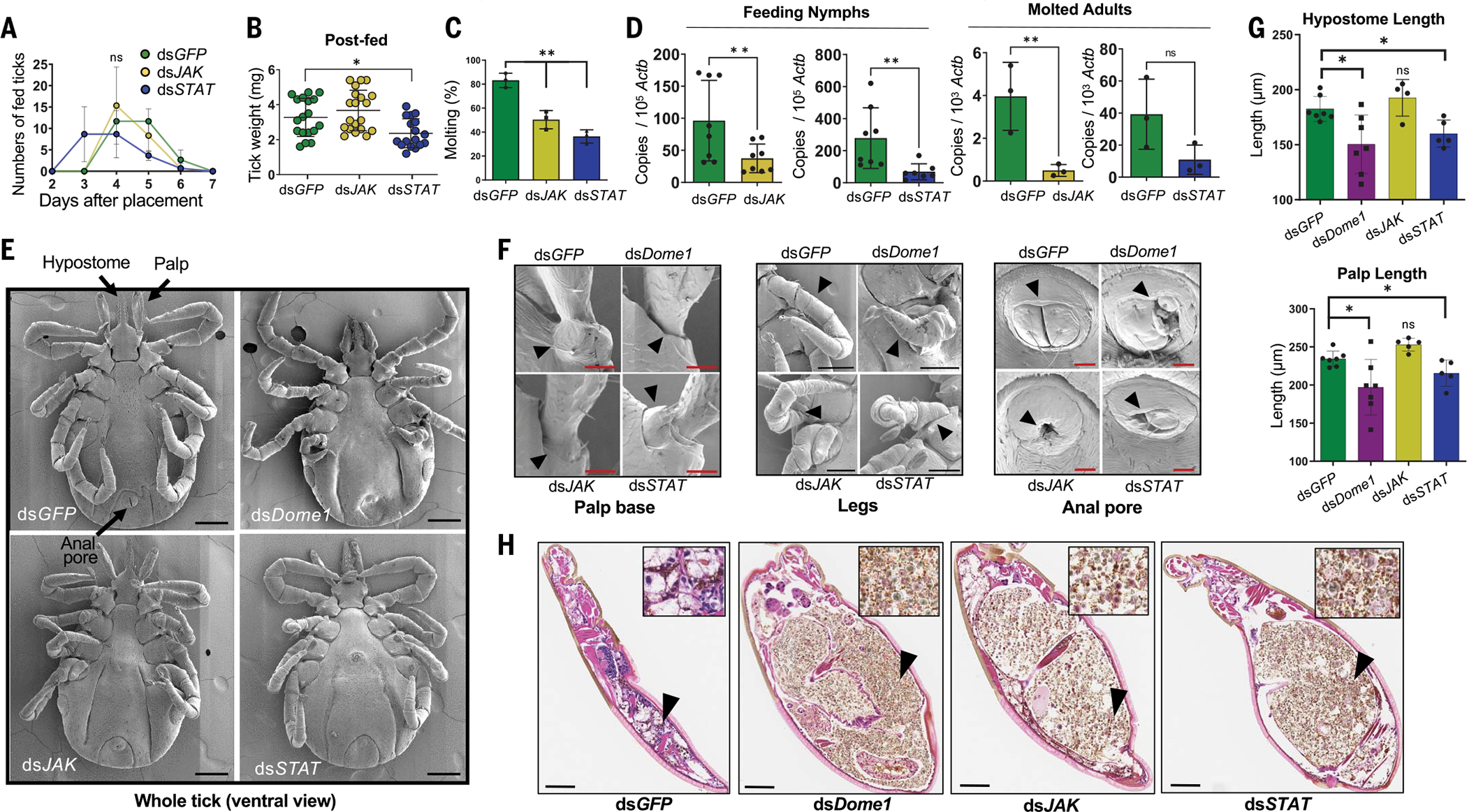
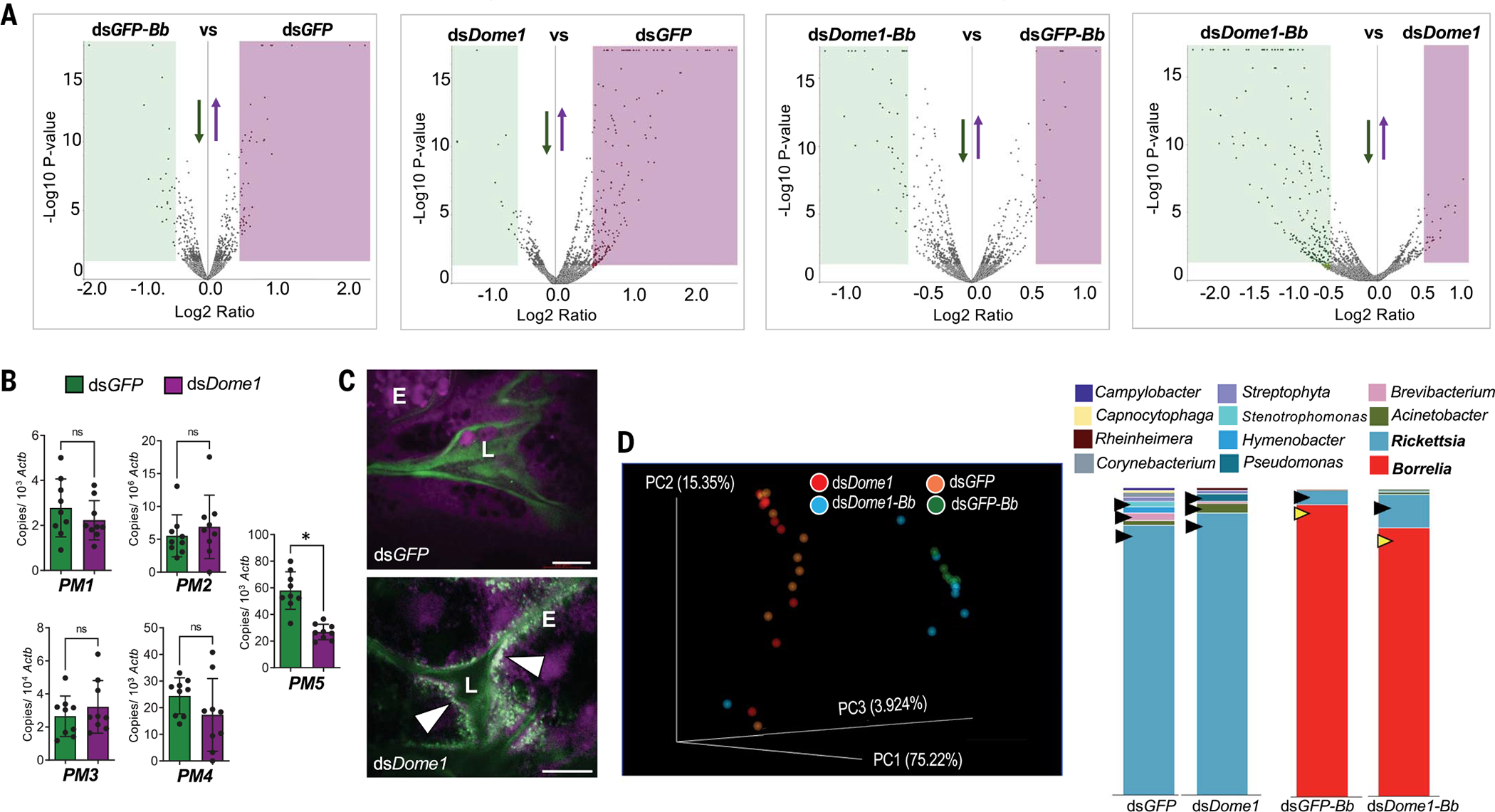
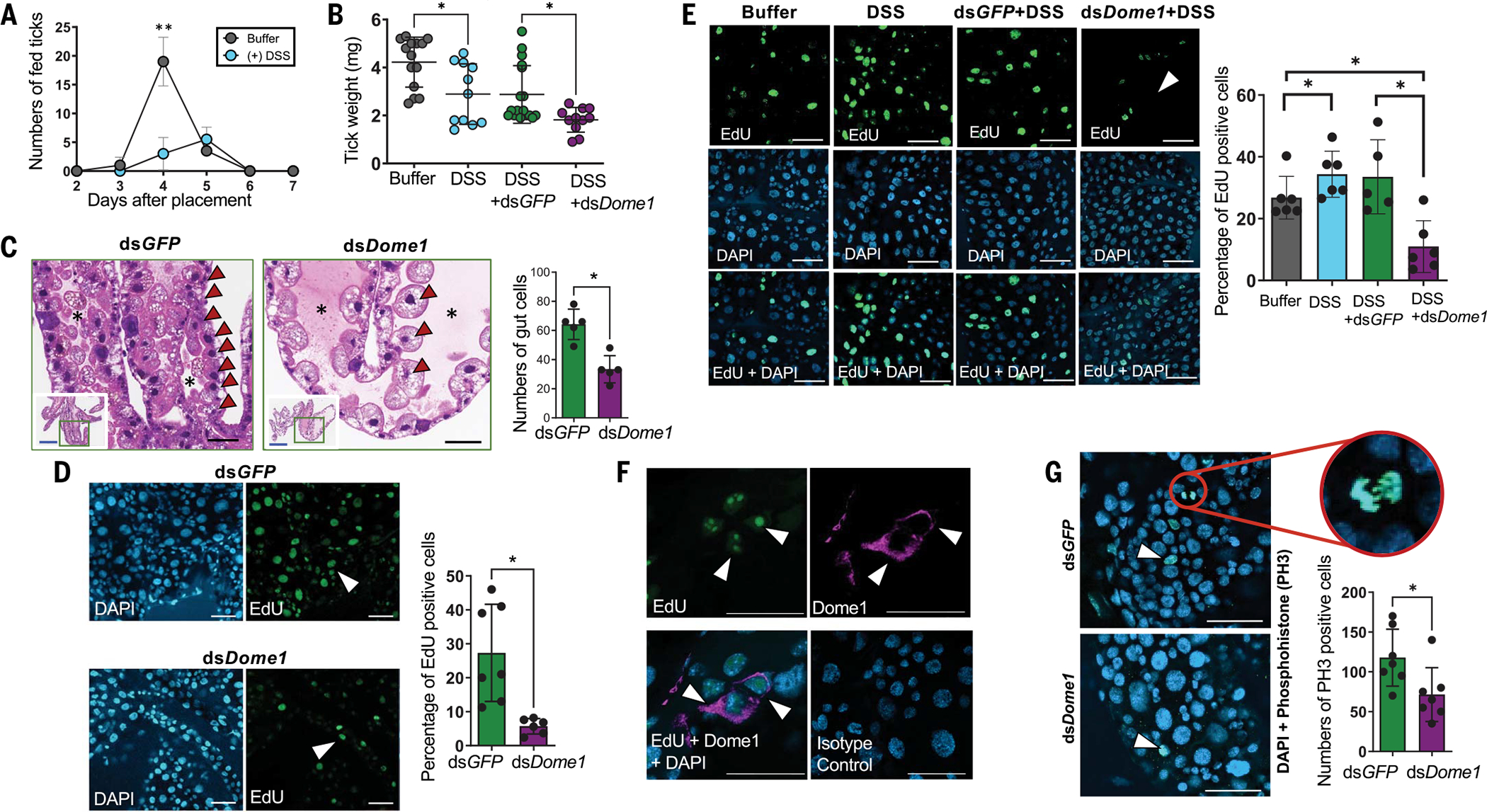
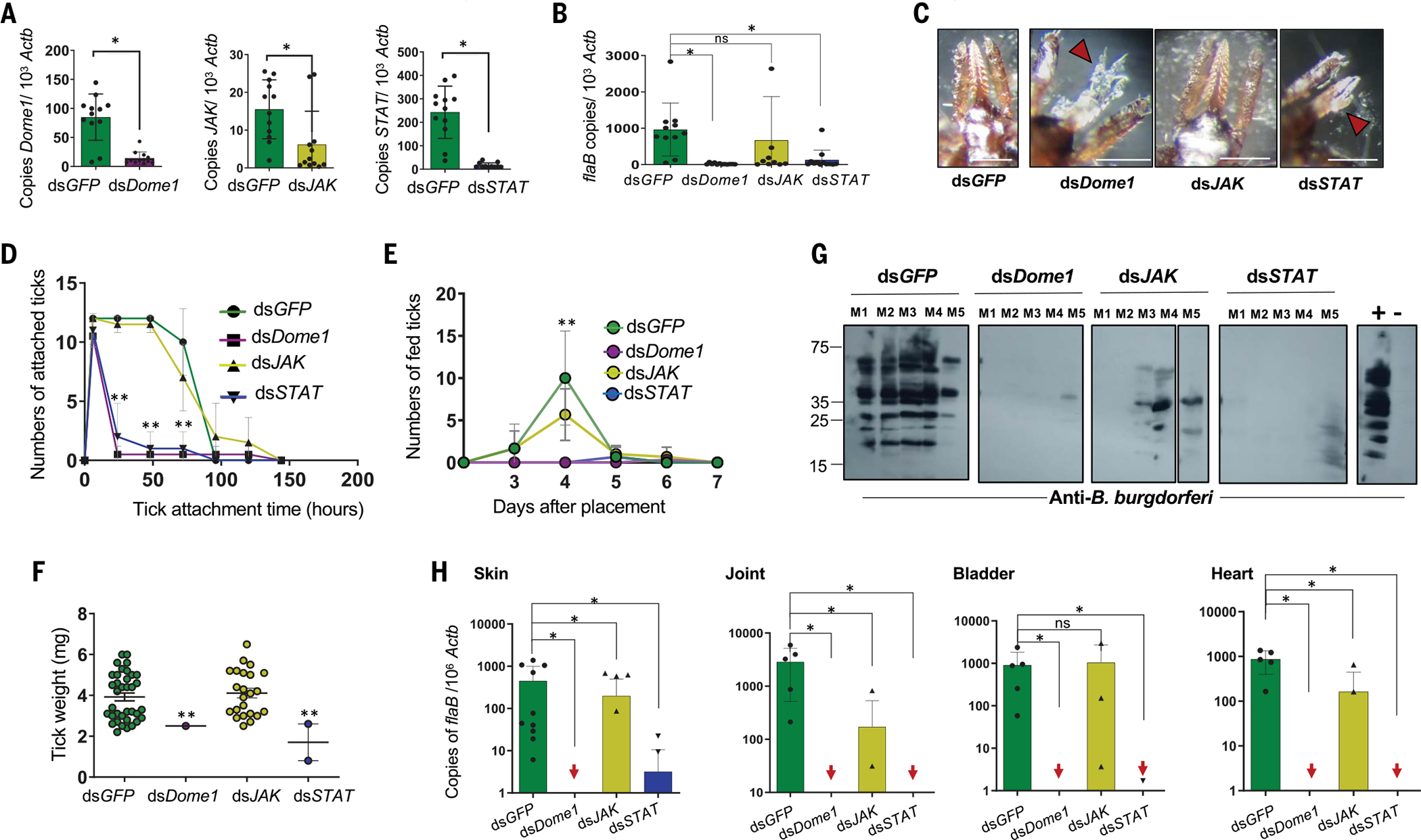
Ancestral signaling pathways serve critical roles in metazoan development, physiology, and immunity. We report an evolutionary interspecies communication pathway involving a central Ixodes scapularis tick receptor termed Dome1, which acquired a mammalian cytokine receptor motif exhibiting high affinity for interferon-gamma (IFN-γ). Host-derived IFN-γ facilitates Dome1-mediated activation of the Ixodes JAK-STAT pathway. This accelerates tick blood meal acquisition and development while upregulating antimicrobial components. The Dome1-JAK-STAT pathway, which exists in most Ixodid tick genomes, regulates the regeneration and proliferation of gut cells-including stem cells-and dictates metamorphosis through the Hedgehog and Notch-Delta networks, ultimately affecting Ixodes vectorial competence. We highlight the evolutionary dependence of I. scapularis on mammalian hosts through cross-species signaling mechanisms that dually influence arthropod immunity and development.
Conflict of interest statement
Figures
References
-
- WHO, Vector-borne diseases. https://www.who.int/mediacentre/factsheets/fs387/en/, (WHO, 2016).
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
