

Palmitoylation couples insulin hypersecretion with β cell failure in diabetes
- PMID: 36634673
- PMCID: PMC9908855
- DOI: 10.1016/j.cmet.2022.12.012
Palmitoylation couples insulin hypersecretion with β cell failure in diabetes
Abstract
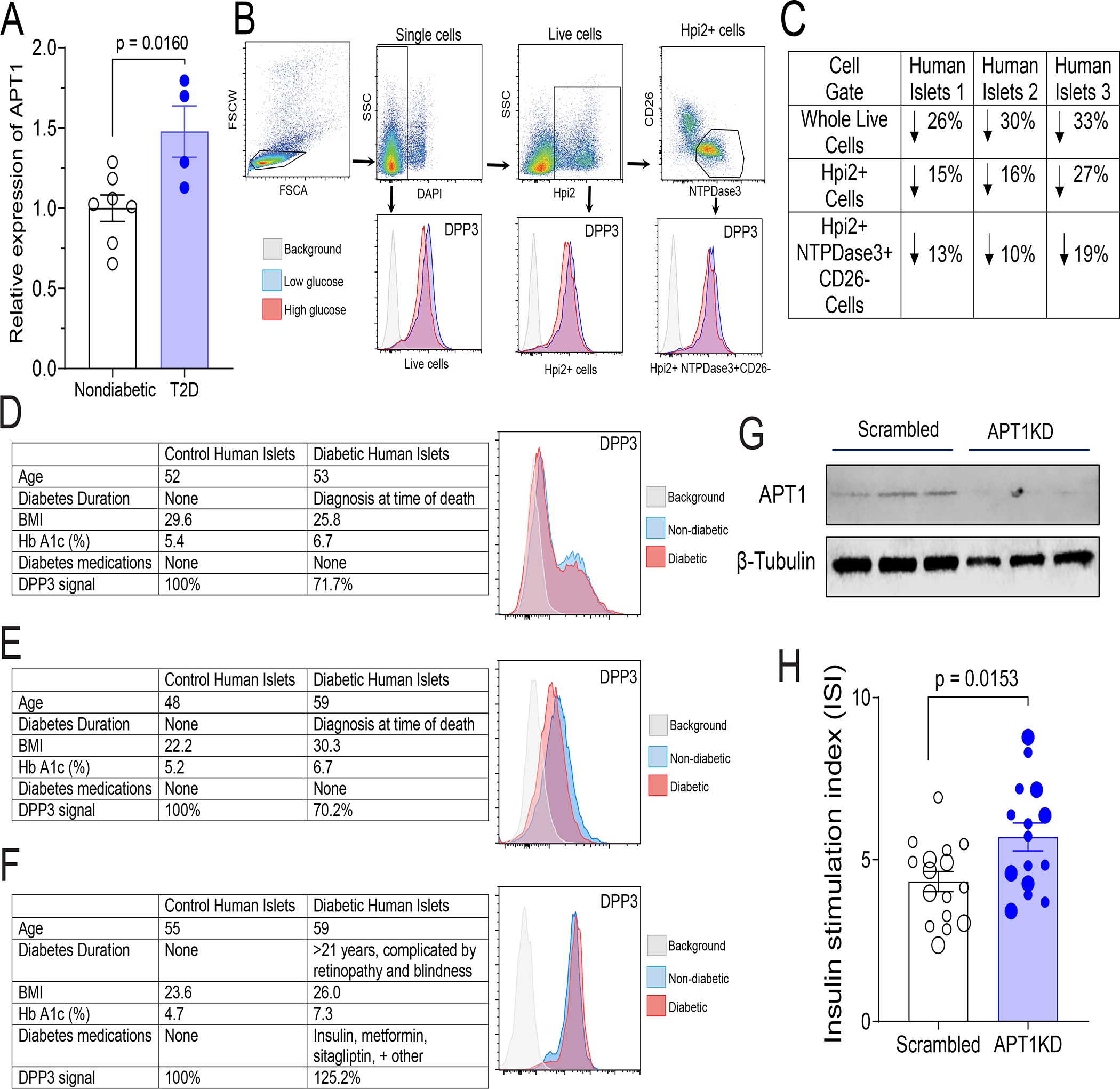
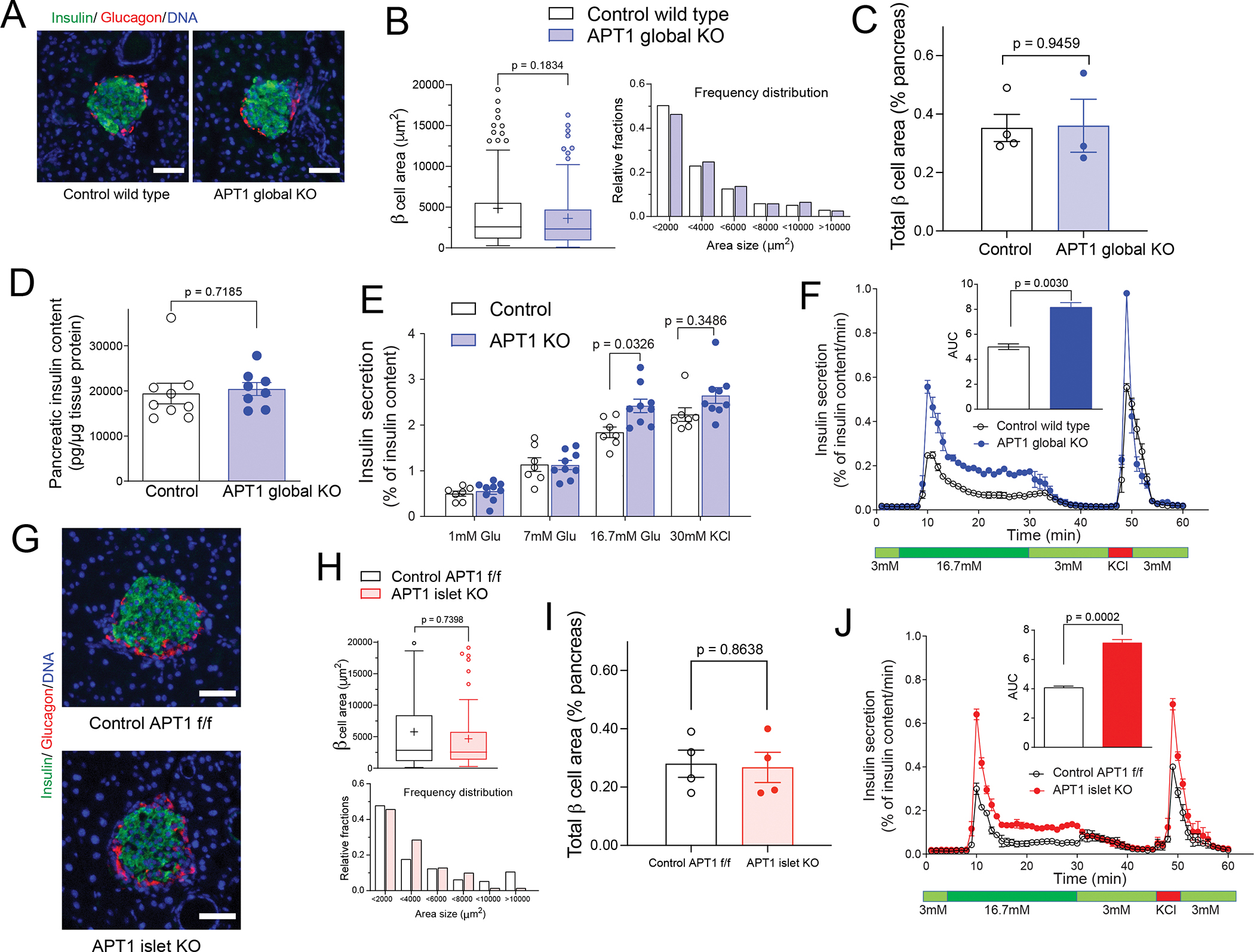
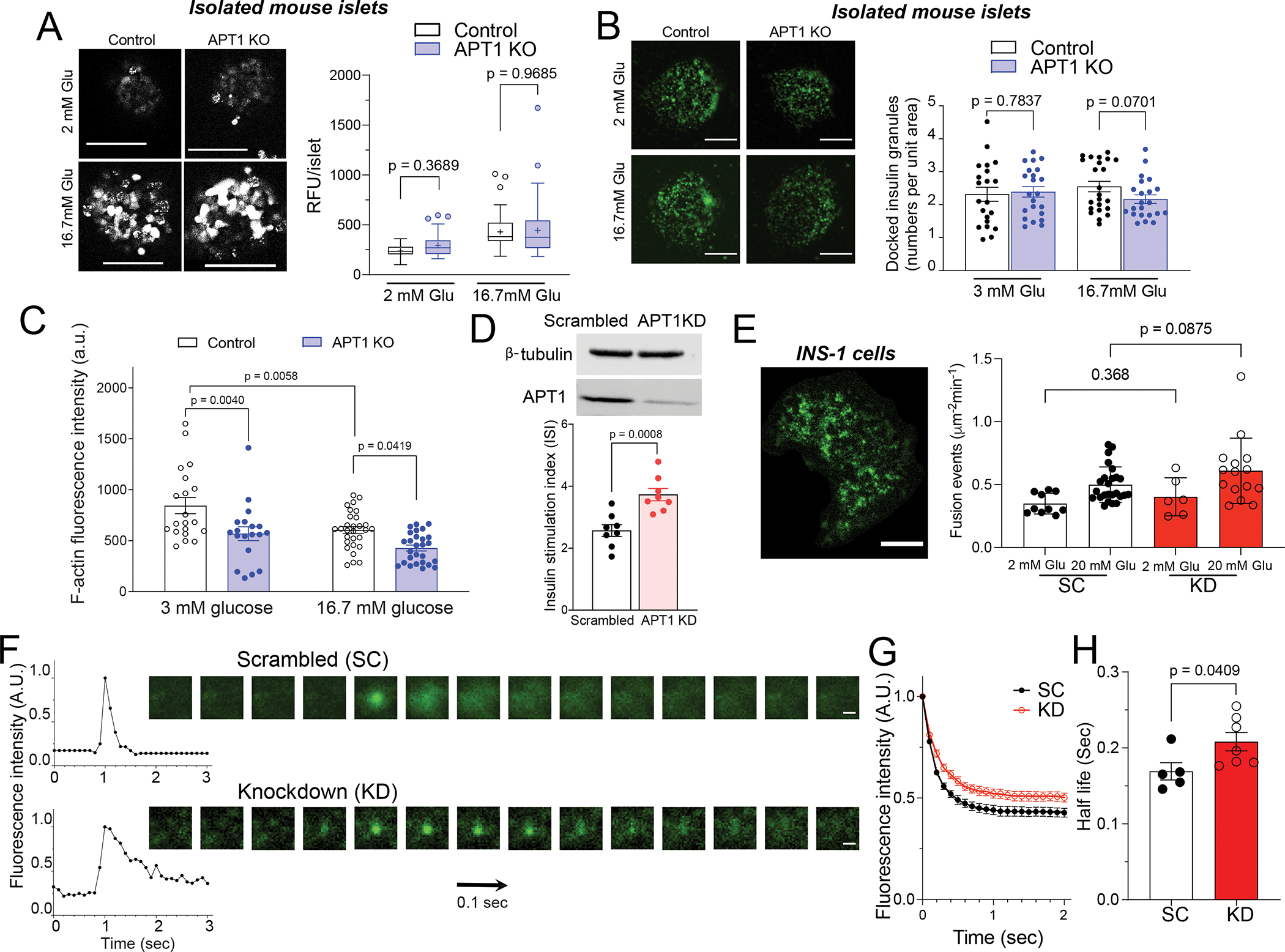
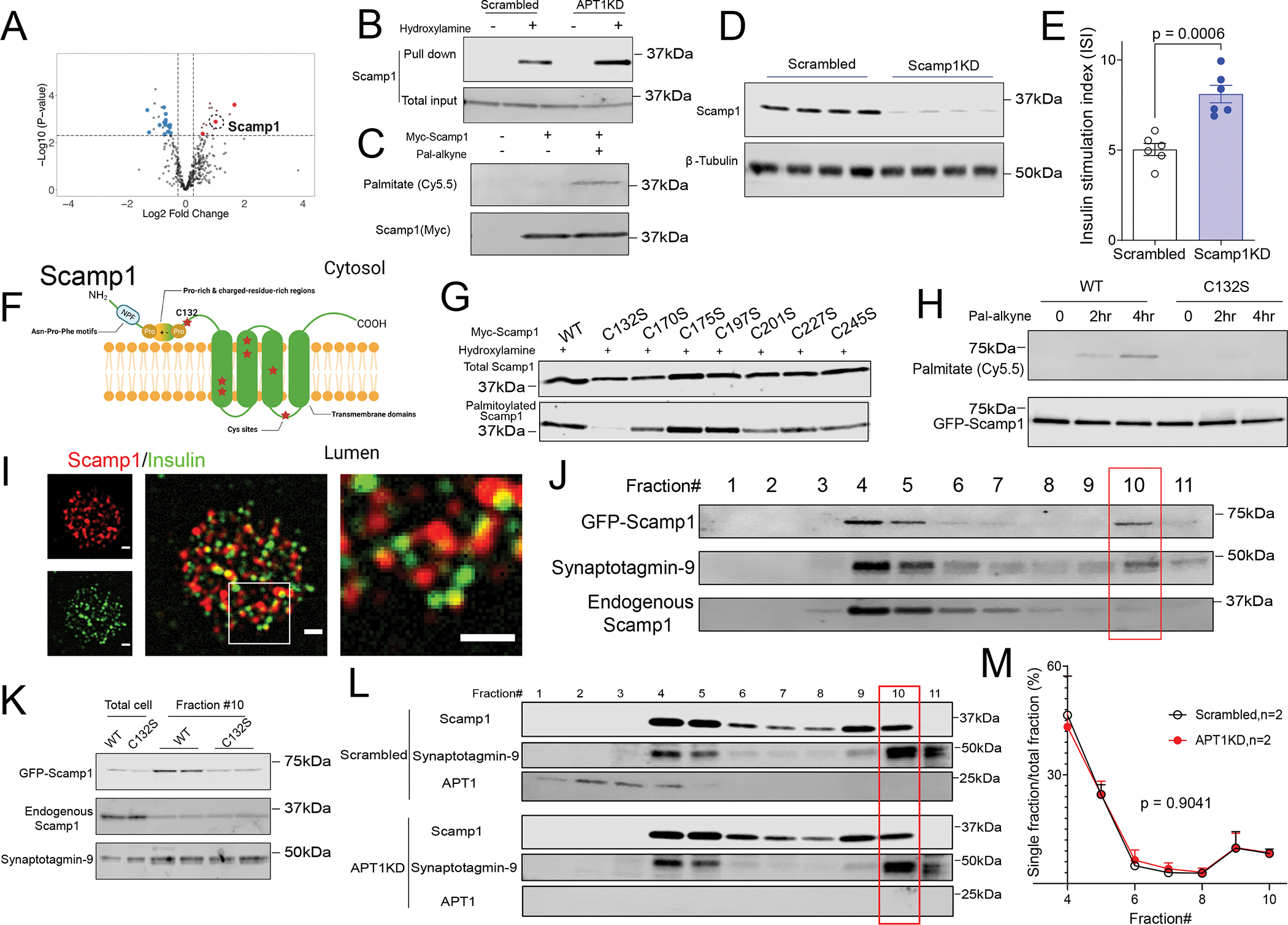
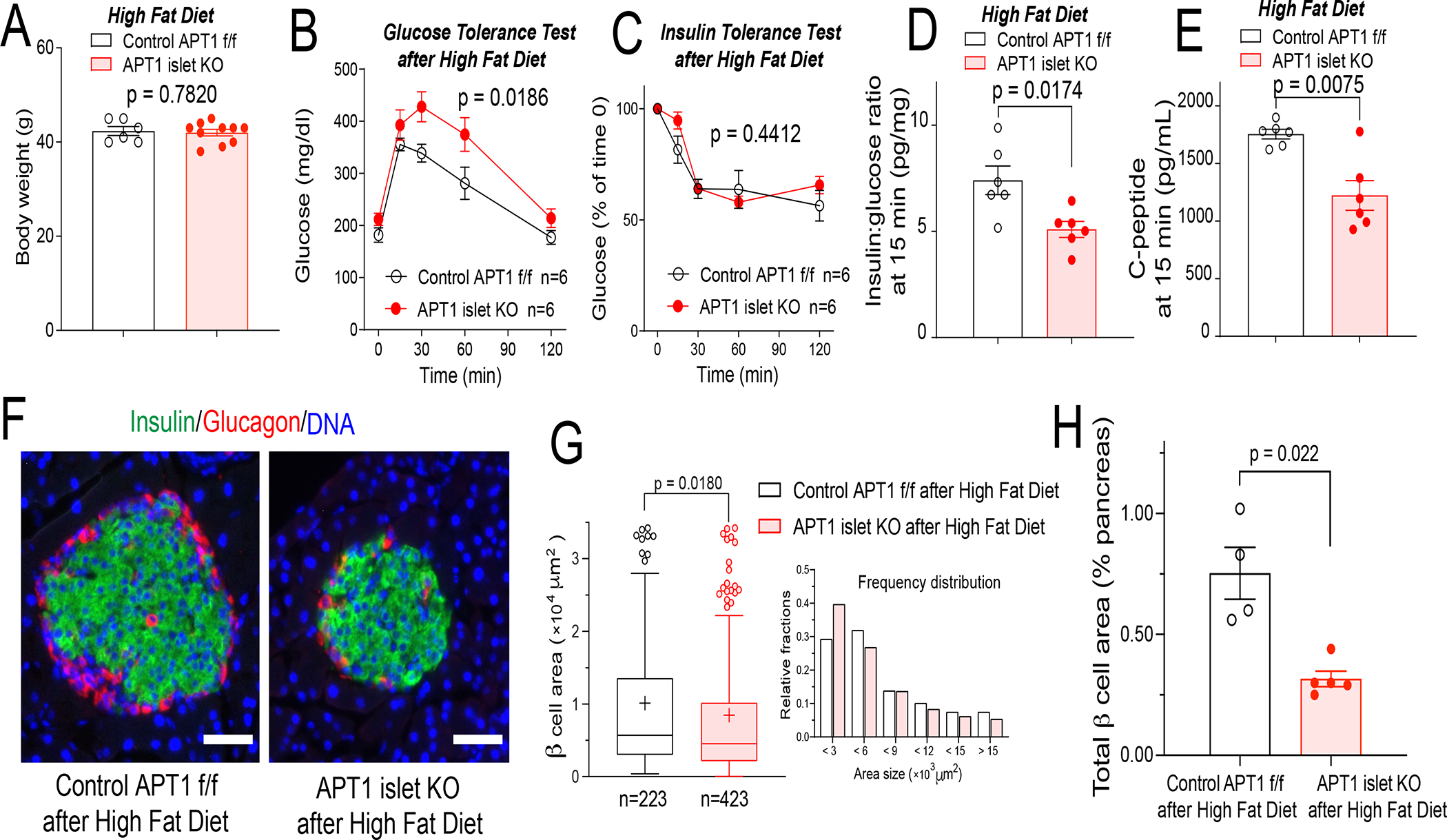
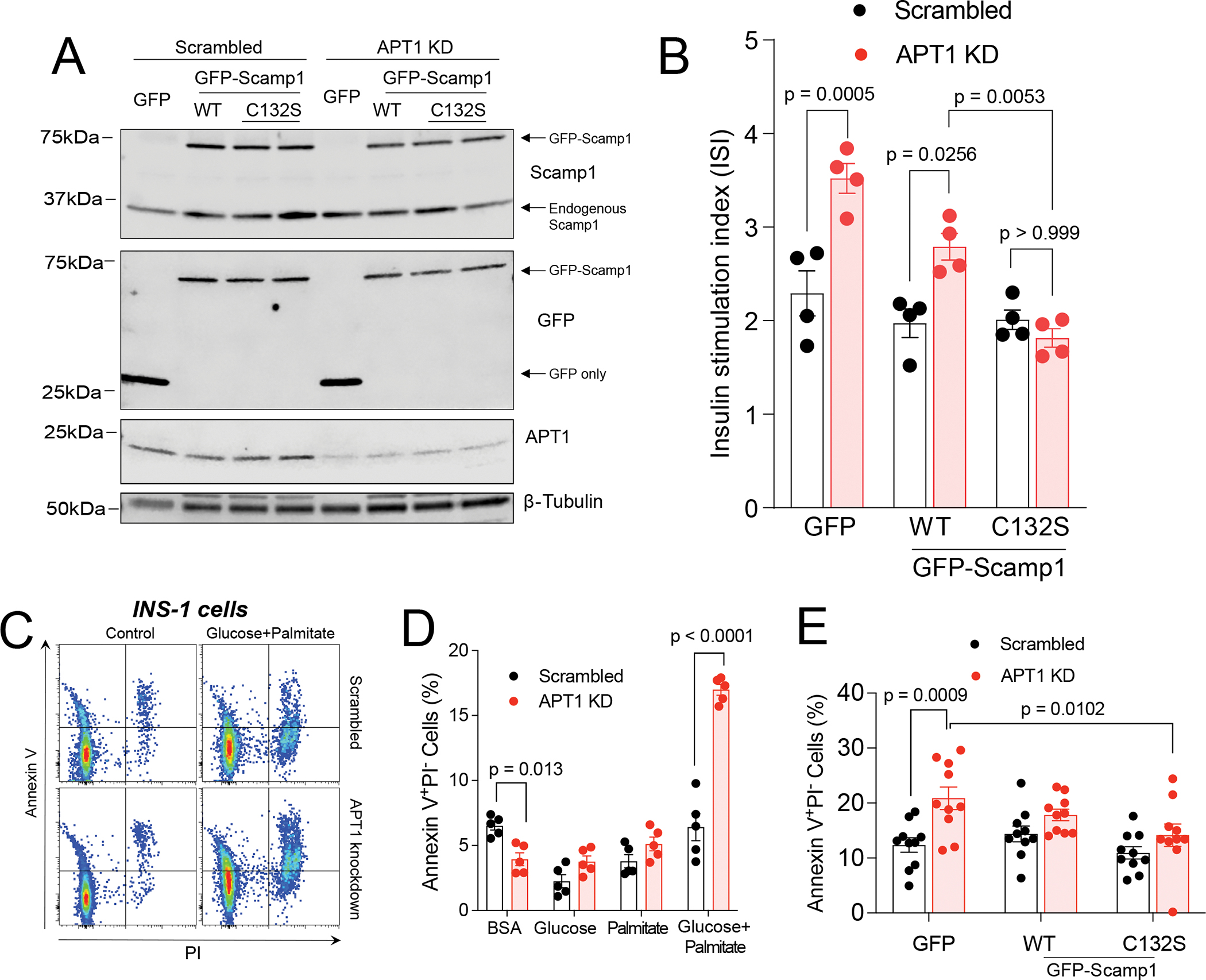
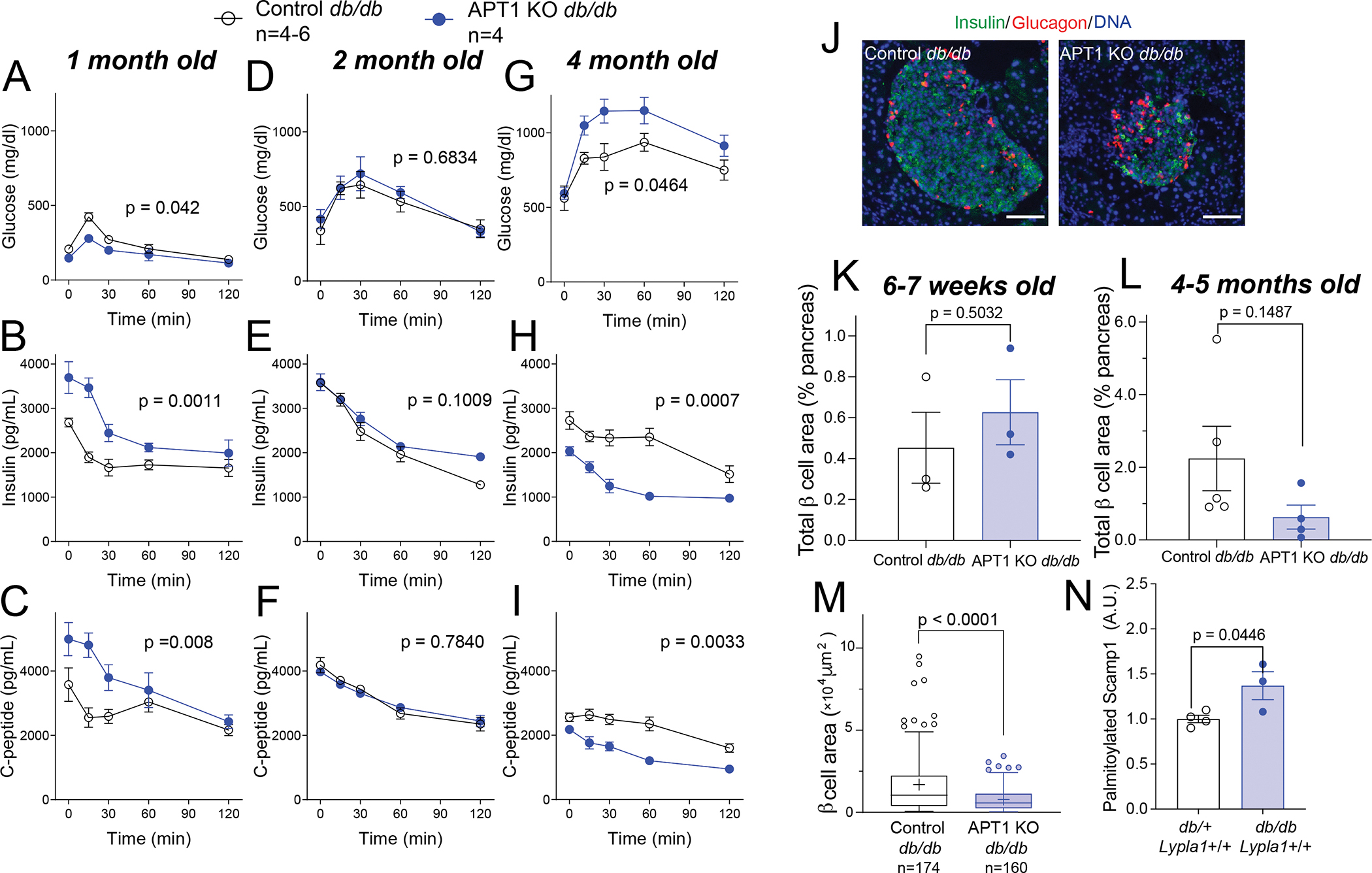
Hyperinsulinemia often precedes type 2 diabetes. Palmitoylation, implicated in exocytosis, is reversed by acyl-protein thioesterase 1 (APT1). APT1 biology was altered in pancreatic islets from humans with type 2 diabetes, and APT1 knockdown in nondiabetic islets caused insulin hypersecretion. APT1 knockout mice had islet autonomous increased glucose-stimulated insulin secretion that was associated with prolonged insulin granule fusion. Using palmitoylation proteomics, we identified Scamp1 as an APT1 substrate that localized to insulin secretory granules. Scamp1 knockdown caused insulin hypersecretion. Expression of a mutated Scamp1 incapable of being palmitoylated in APT1-deficient cells rescued insulin hypersecretion and nutrient-induced apoptosis. High-fat-fed islet-specific APT1-knockout mice and global APT1-deficient db/db mice showed increased β cell failure. These findings suggest that APT1 is regulated in human islets and that APT1 deficiency causes insulin hypersecretion leading to β cell failure, modeling the evolution of some forms of human type 2 diabetes.
Keywords: S-acylation; acyl-protein thioesterases; beta cell failure; lipotoxicity; type 2 diabetes.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests C.F.S. is a member of the Cell Metabolism advisory board.
Figures
References
-
- Tura A, Grespan E, Gobl CS, Koivula RW, Franks PW, Pearson ER, Walker M, Forgie IM, Giordano GN, Pavo I, et al. (2021). Profiles of Glucose Metabolism in Different Prediabetes Phenotypes, Classified by Fasting Glycemia, 2-Hour OGTT, Glycated Hemoglobin, and 1-Hour OGTT: An IMI DIRECT Study. Diabetes 70, 2092–2106. 10.2337/db21-0227. - DOI - PubMed
-
- Weyer C, Hanson RL, Tataranni PA, Bogardus C, and Pratley RE (2000). A high fasting plasma insulin concentration predicts type 2 diabetes independent of insulin resistance: evidence for a pathogenic role of relative hyperinsulinemia. Diabetes 49, 2094–2101. 10.2337/diabetes.49.12.2094. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R35 GM119840/GM/NIGMS NIH HHS/United States
- R01 DK115972/DK/NIDDK NIH HHS/United States
- R01 DK132090/DK/NIDDK NIH HHS/United States
- P30 DK056341/DK/NIDDK NIH HHS/United States
- R01 DK101392/DK/NIDDK NIH HHS/United States
- R01 HL157154/HL/NHLBI NIH HHS/United States
- T32 HL125241/HL/NHLBI NIH HHS/United States
- P30 DK020579/DK/NIDDK NIH HHS/United States
- P60 DK020579/DK/NIDDK NIH HHS/United States
- R01 DK123301/DK/NIDDK NIH HHS/United States
- R01 DK112921/DK/NIDDK NIH HHS/United States
- F30 DK131830/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
