

F4/80+Ly6Chigh Macrophages Lead to Cell Plasticity and Cancer Initiation in Colitis
- PMID: 36634827
- PMCID: PMC10038892
- DOI: 10.1053/j.gastro.2023.01.002
F4/80+Ly6Chigh Macrophages Lead to Cell Plasticity and Cancer Initiation in Colitis
Abstract
Background & aims: Colorectal cancer is a leading cause of cancer death, and a major risk factor is chronic inflammation. Despite the link between colitis and cancer, the mechanism by which inflammation leads to colorectal cancer is not well understood.
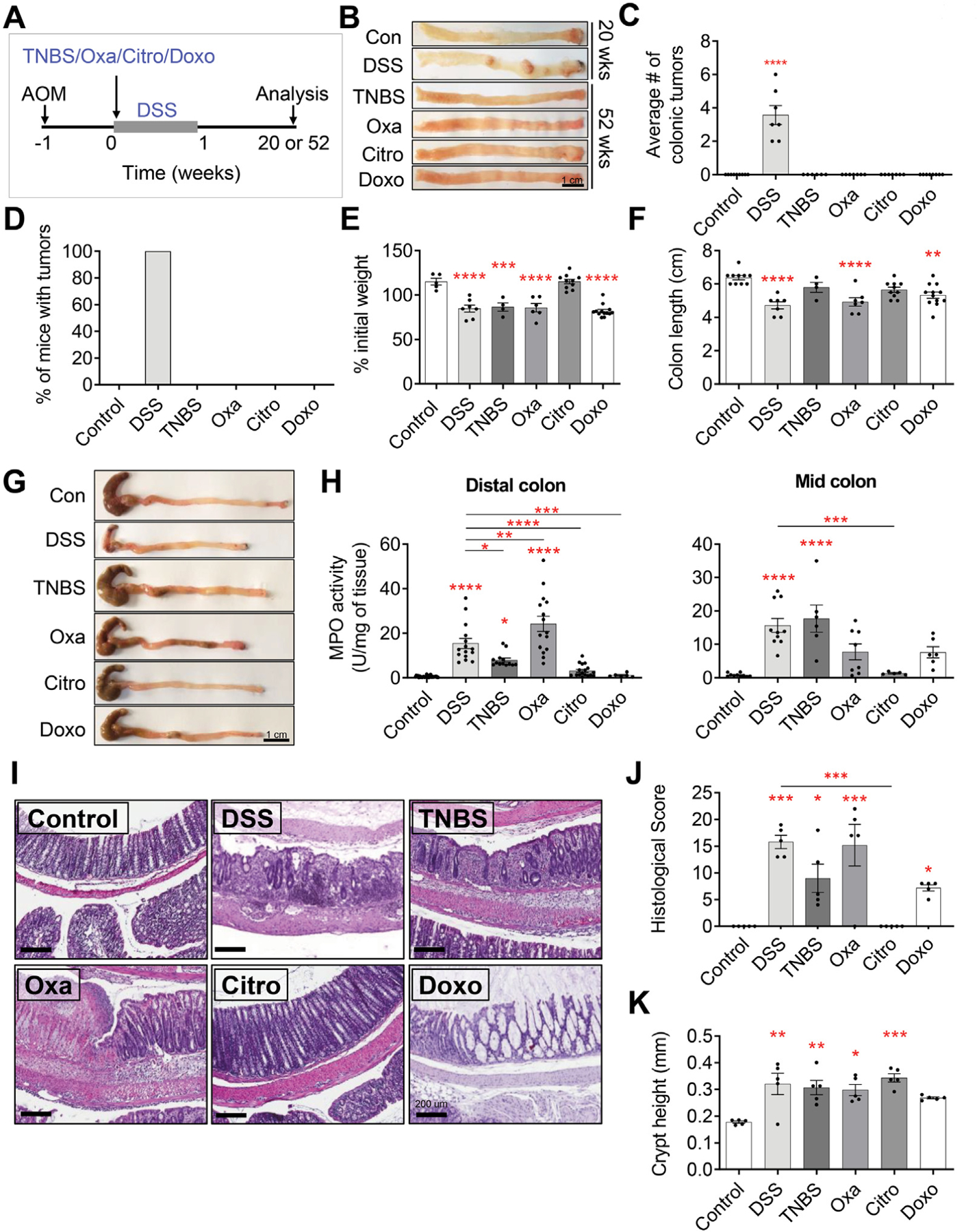
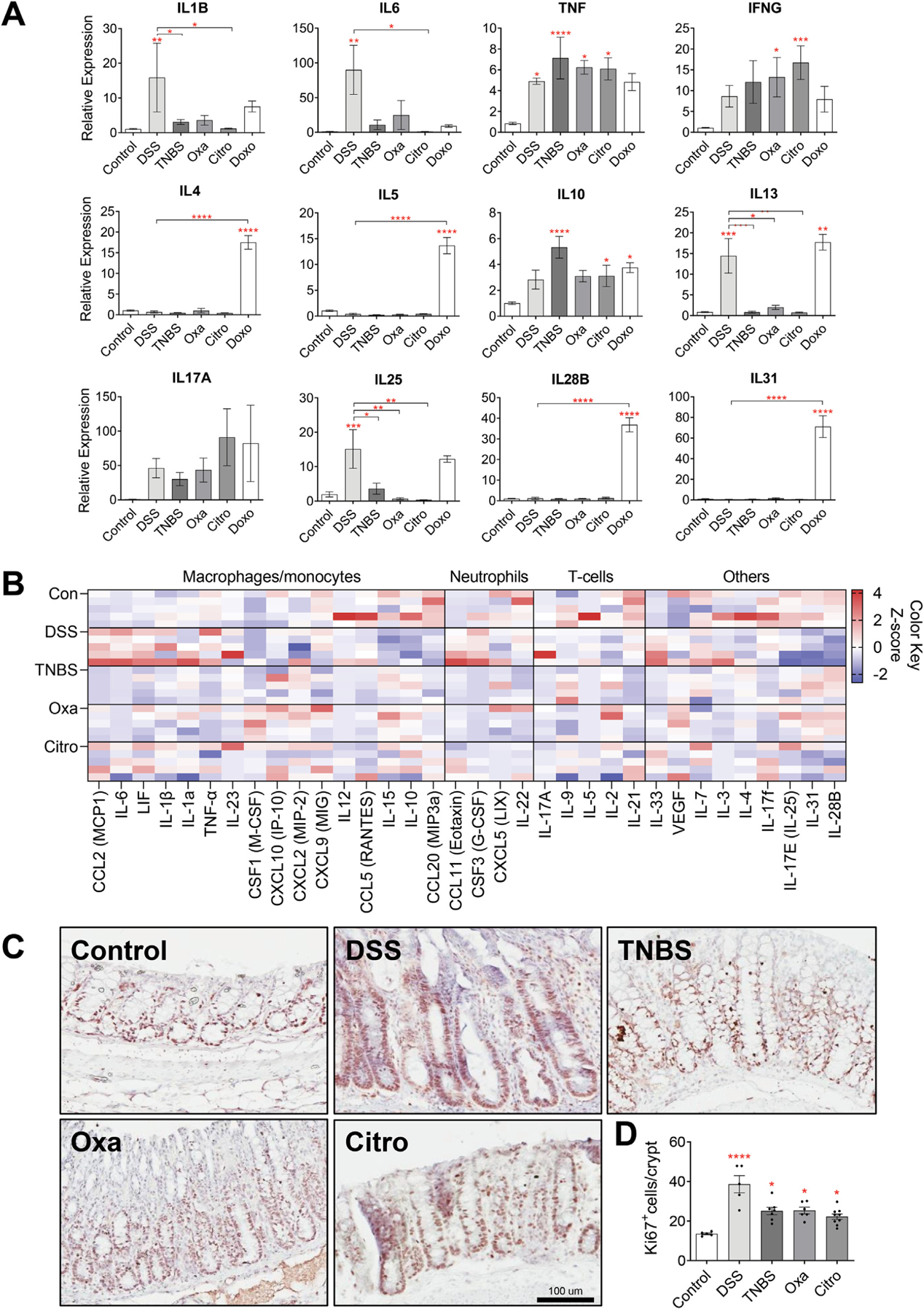
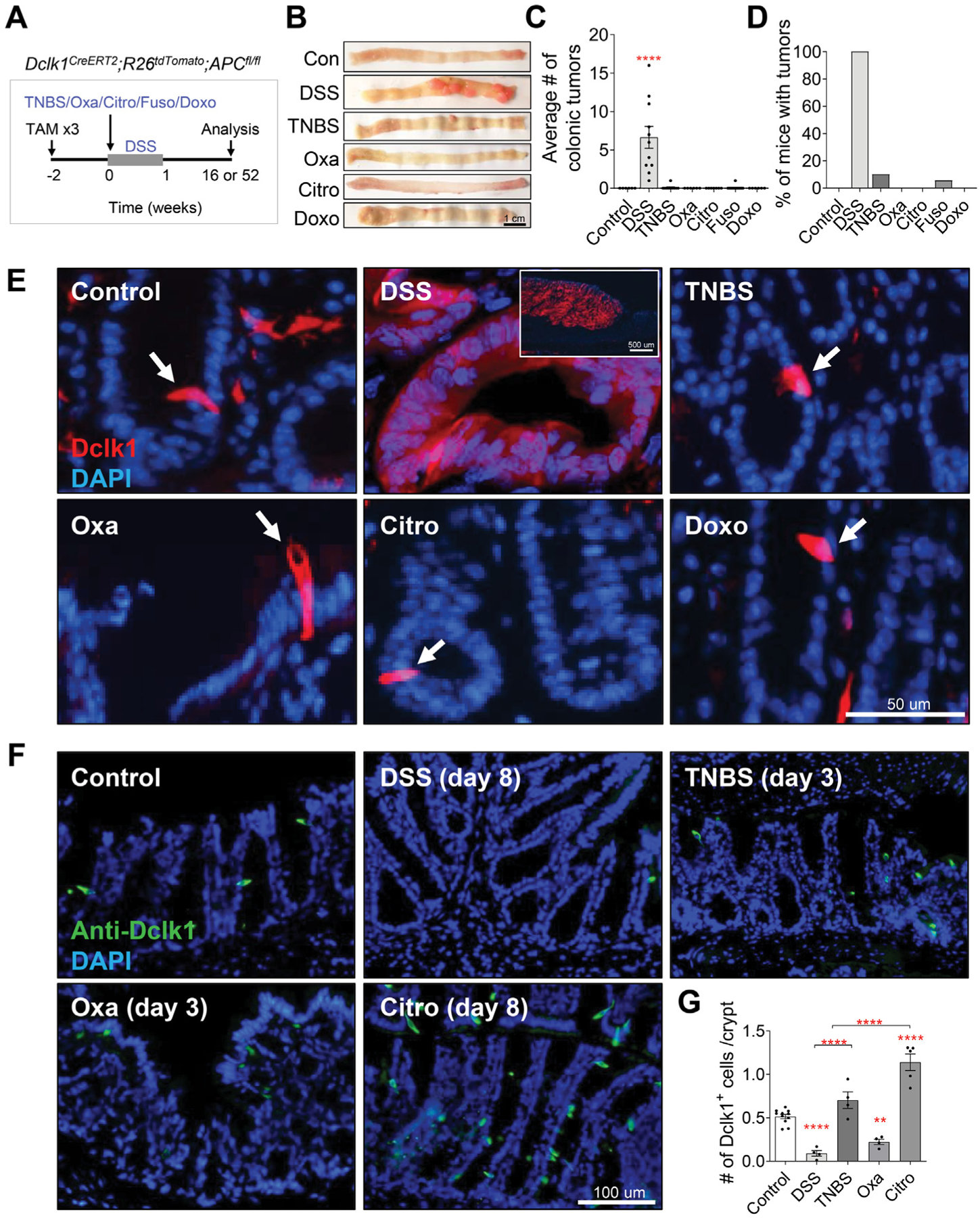
Methods: To investigate whether different forms of inflammation pose the same risk of cancer, we compared several murine models of colitis (dextran sodium sulfate [DSS], 2,4,6-trinitrobenzene sulfonic acid, 4-ethoxylmethylene-2-phenyloxazol-5-one, Citrobacter rodentium, Fusobacterium nucleatum, and doxorubicin) with respect to their ability to lead to colonic tumorigenesis. We attempted to correlate the severity of colitis and inflammatory profile with the risk of tumorigenesis in both azoxymethane-dependent and Dclk1/APCfl/fl murine models of colitis-associated cancer.
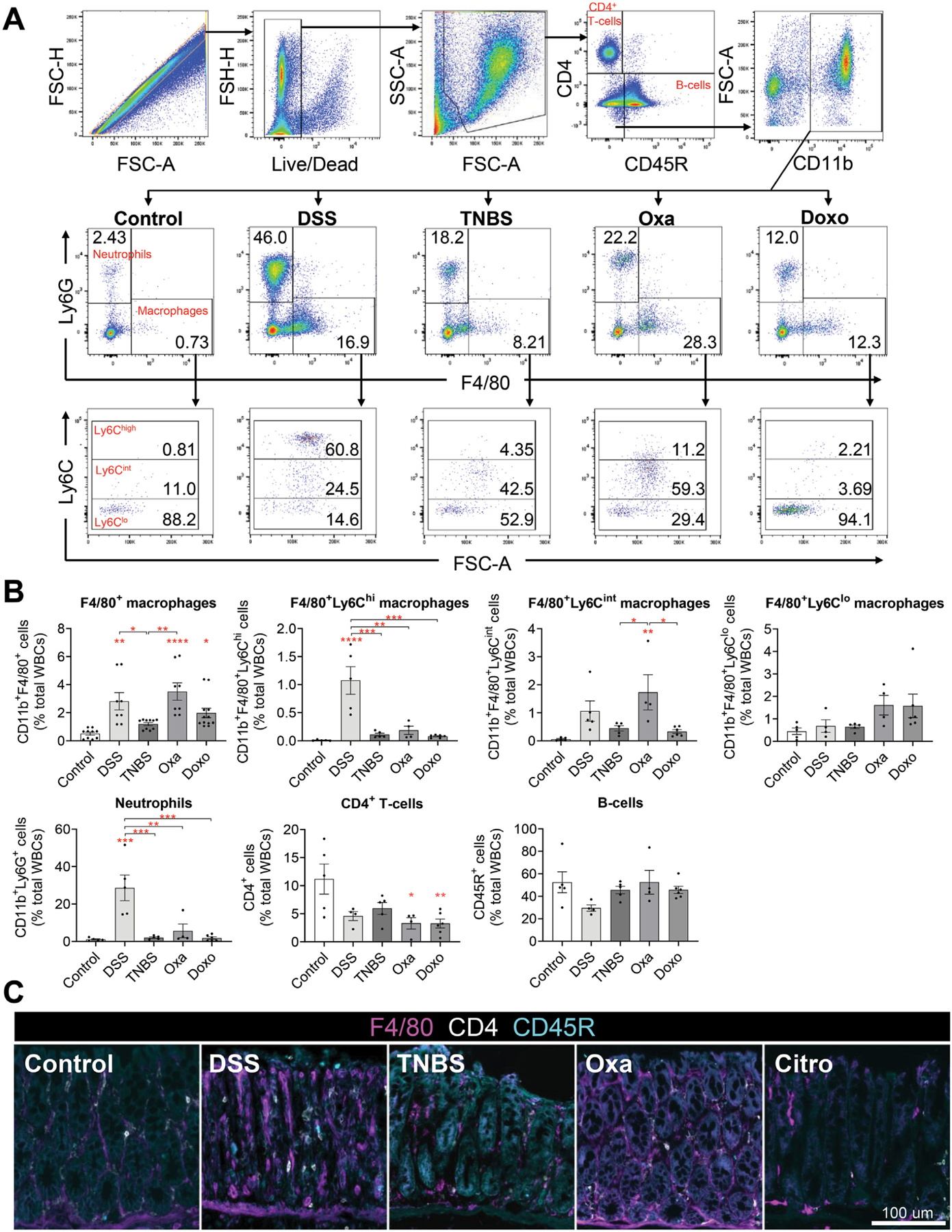
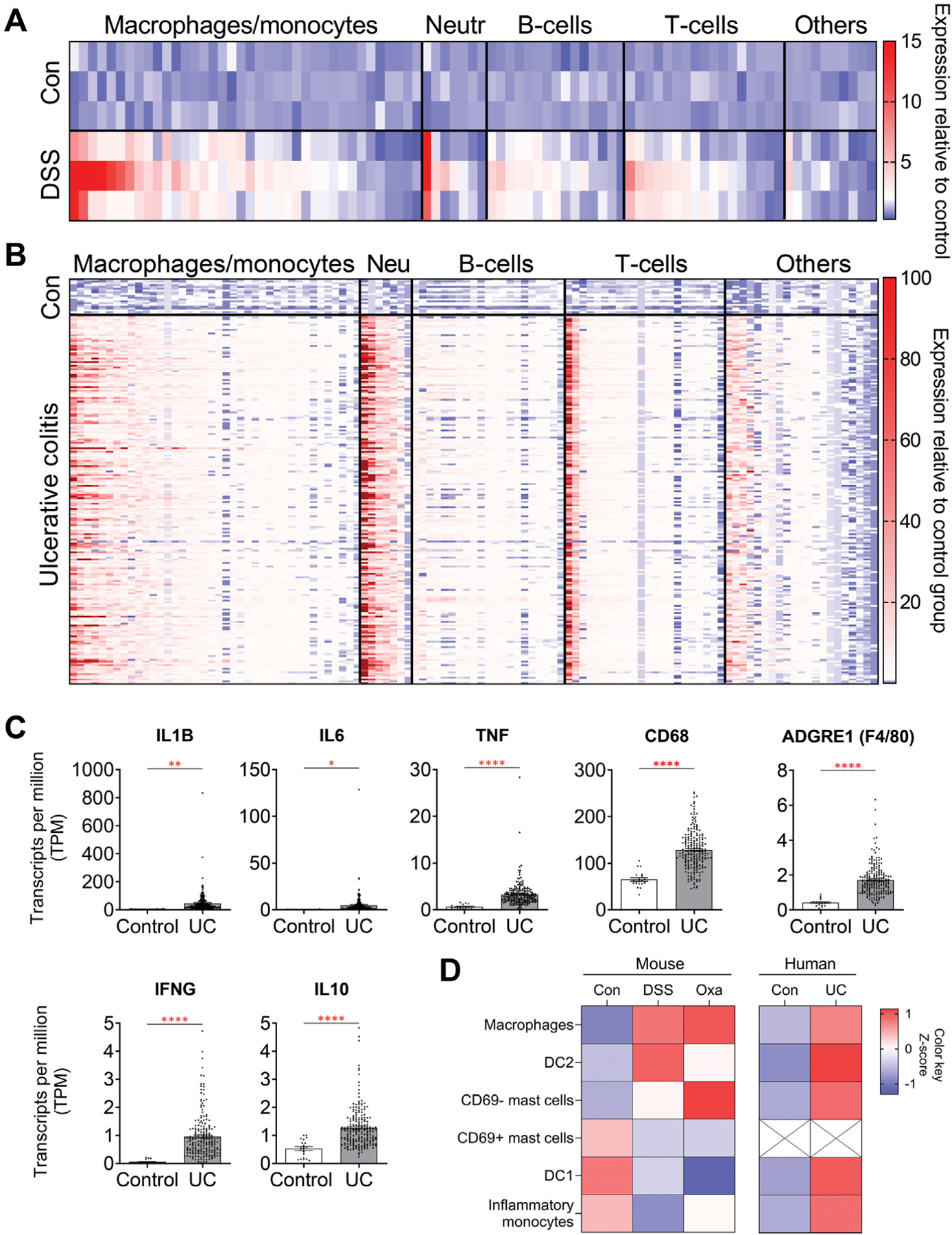
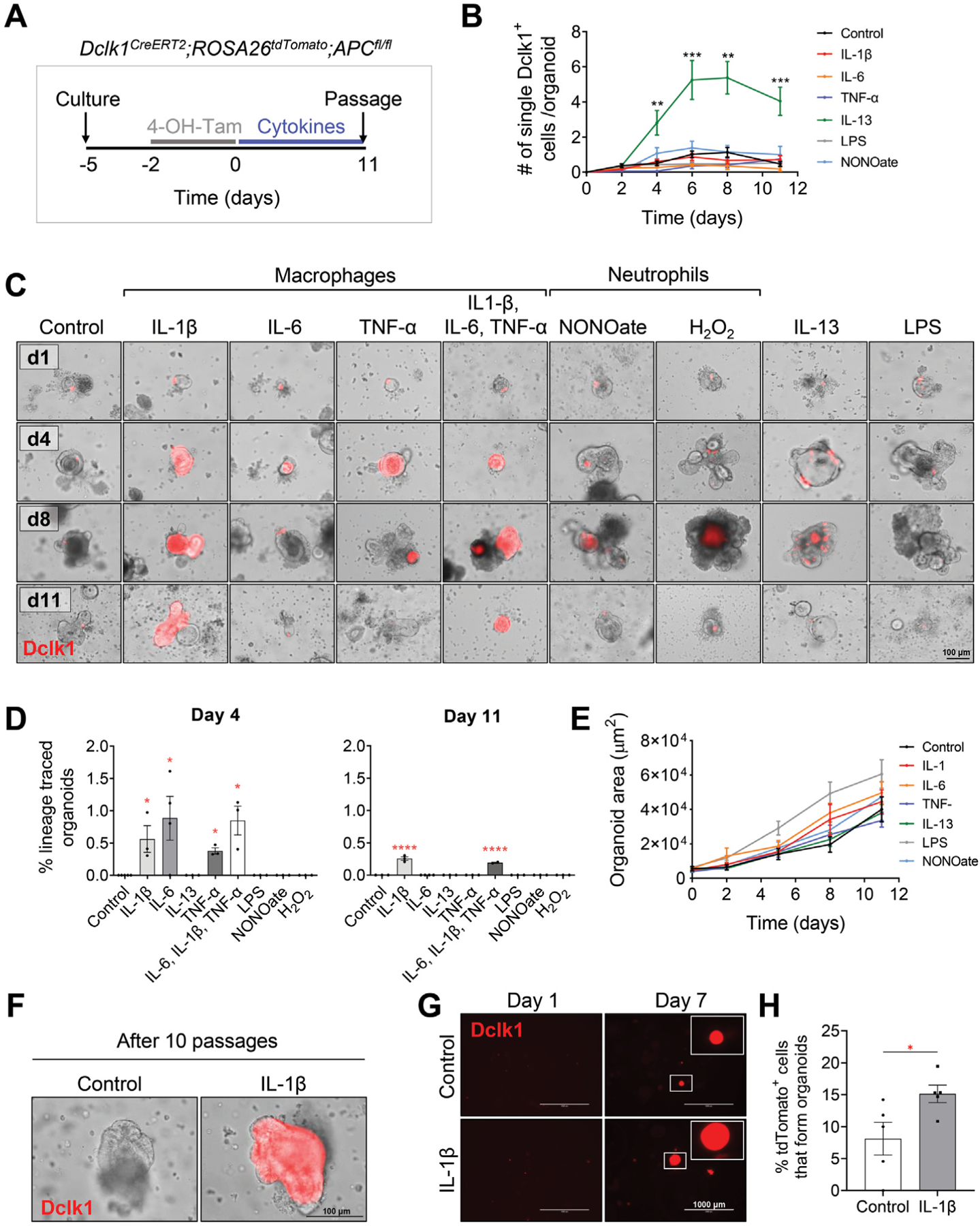
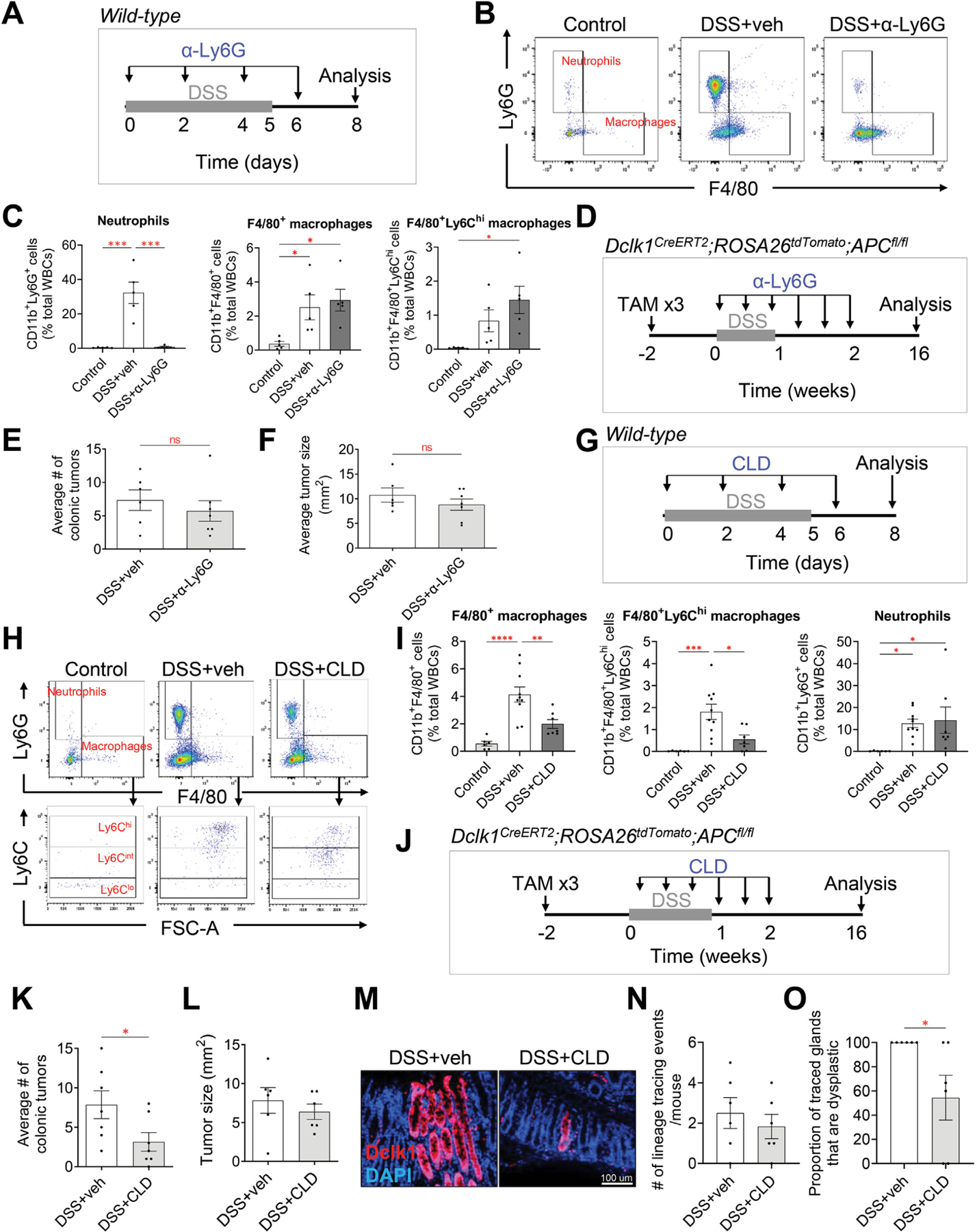
Results: DSS colitis reproducibly led to colonic tumors in both mouse models of colitis-associated cancer. In contrast, all other forms of colitis did not lead to cancer. When compared with the colitis not associated with tumorigenesis, DSS colitis was characterized by significantly increased CD11b+F4/80+Ly6Chigh macrophages and CD11b+Ly6G+ neutrophils. Interestingly, depletion of the CD11b+F4/80+Ly6Chigh macrophages inhibited tumorigenesis, whereas depletion of CD11b+Ly6G+ neutrophils had no effect on tumorigenesis. Furthermore, the macrophage-derived cytokines interleukin-1β, tumor necrosis factor-α, and interleukin-6 were significantly increased in DSS colitis and promoted stemness of Dclk1+ tuft cells that serve as the cellular origin of cancer.
Conclusions: We have identified CD11b+F4/80+Ly6Chigh macrophages as key mediators of cancer initiation in colitis-associated cancer. Development of new therapies that target these cells may provide an effective preventative strategy for colitis-associated cancer.
Keywords: AOM/DSS; Colitis; Colitis-Associated Cancer; Colorectal Cancer; Dclk1; Macrophages; Tuft Cells.
Copyright © 2023 AGA Institute. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest
The authors disclose no conflicts.
Figures
References
-
- GLOBOCAN 2020. Colorectal Cancer. International Agency for Research on Cancer, World Health Organization. Accessed January 25, 2023. https://gco.iarc.fr/today/data/factsheets/cancers/10_8_9-Colorectum-fact....
-
- Ullman TA, Itzkowitz SH. Intestinal inflammation and cancer. Gastroenterology 2011;140:1807–1816. - PubMed
-
- Neufert C, Becker C, Neurath MF. An inducible mouse model of colon carcinogenesis for the analysis of sporadic and inflammation-driven tumor progression. Nat Protoc 2007;2:1998–2004. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
