

STRA6 is essential for induction of vascular smooth muscle lineages in human embryonic cardiac outflow tract development
- PMID: 36635482
- PMCID: PMC10202647
- DOI: 10.1093/cvr/cvad010
STRA6 is essential for induction of vascular smooth muscle lineages in human embryonic cardiac outflow tract development
Abstract
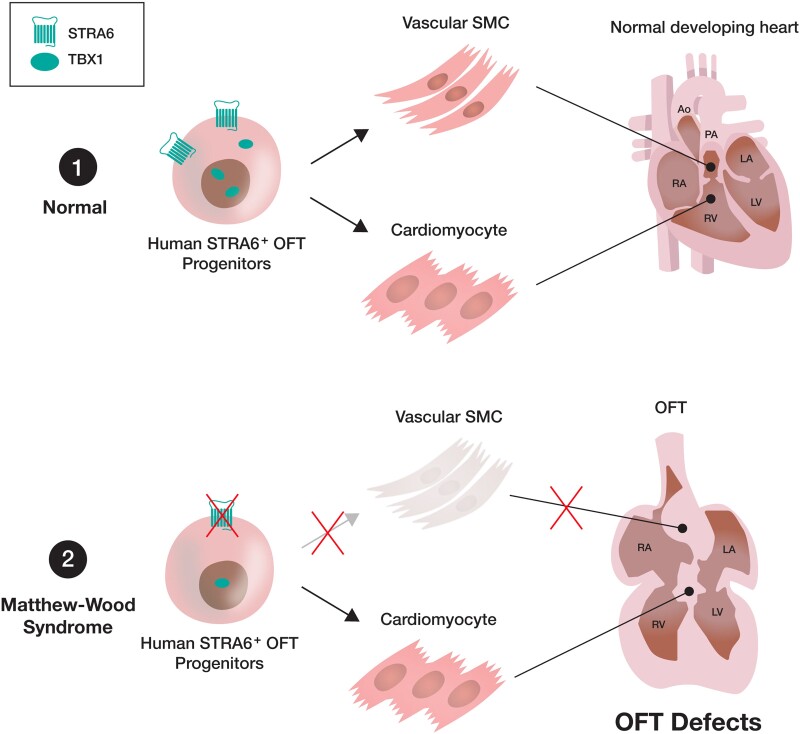
Aims: Retinoic acid (RA) signalling is essential for heart development, and dysregulation of the RA signalling can cause several types of cardiac outflow tract (OFT) defects, the most frequent congenital heart disease (CHD) in humans. Matthew-Wood syndrome is caused by inactivating mutations of a transmembrane protein gene STRA6 that transports vitamin A (retinol) from extracellular into intracellular spaces. This syndrome shows a broad spectrum of malformations including CHD, although murine Stra6-null neonates did not exhibit overt heart defects. Thus, the detailed mechanisms by which STRA6 mutations could lead to cardiac malformations in humans remain unclear. Here, we investigated the role of STRA6 in the context of human cardiogenesis and CHD.
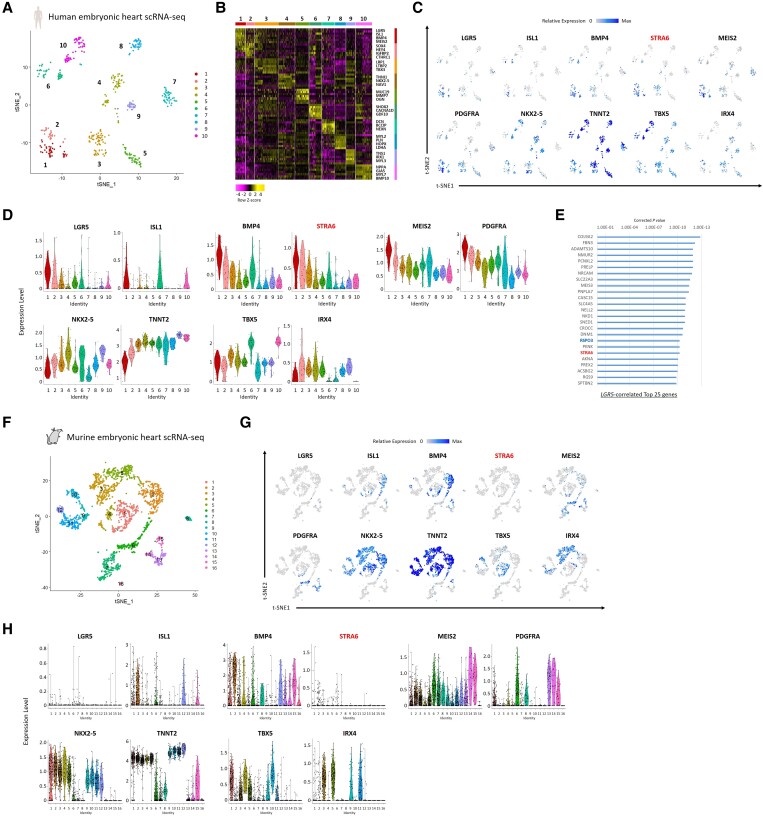
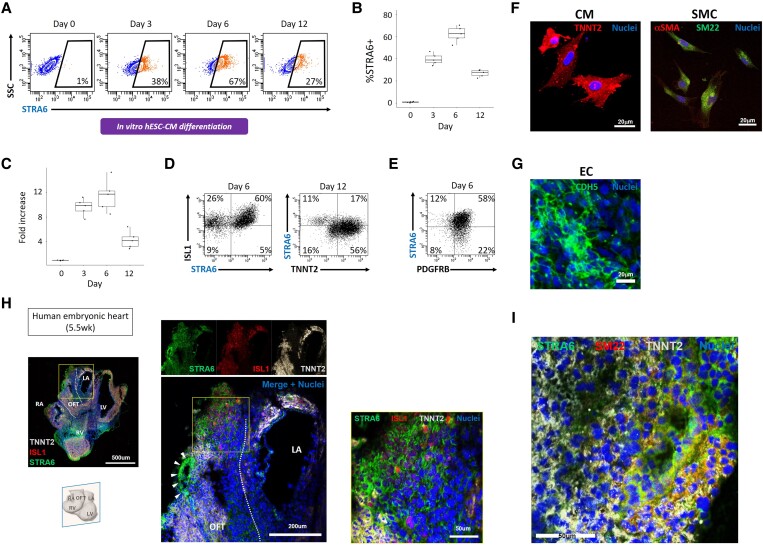
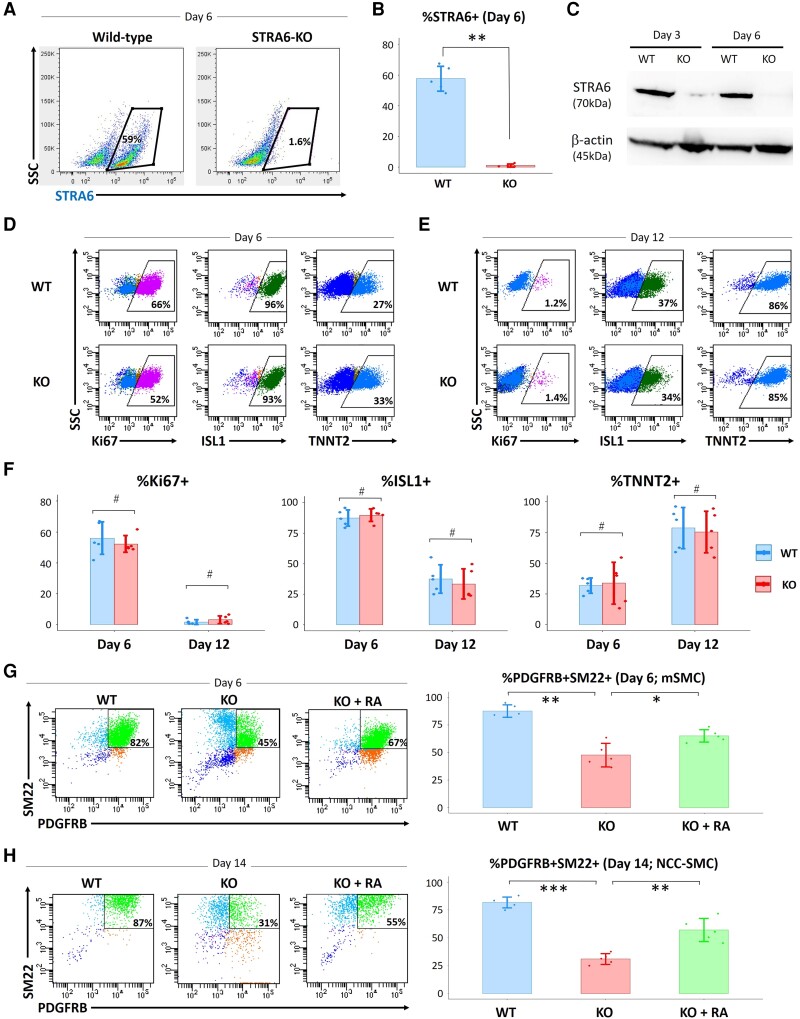
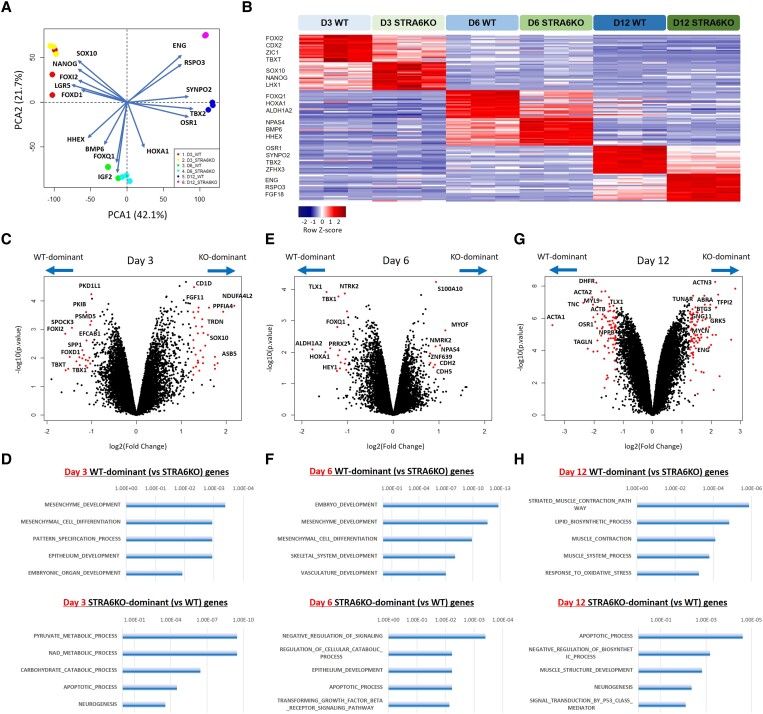
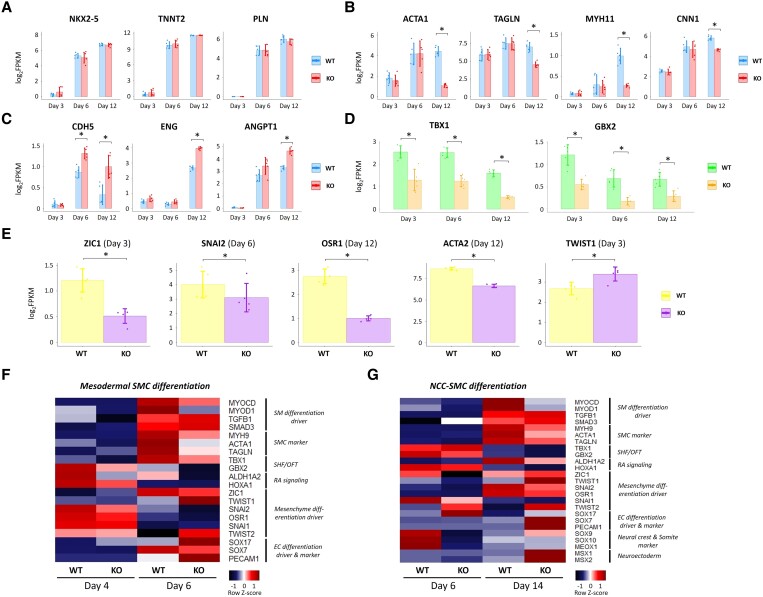
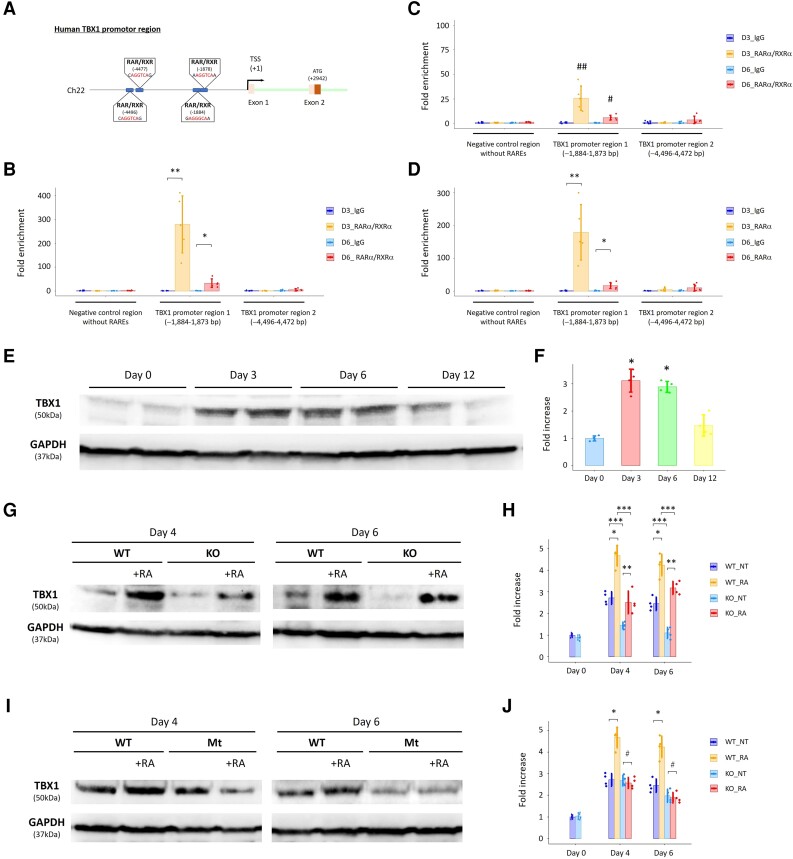
Methods and results: To gain molecular signatures in species-specific cardiac development, we first compared single-cell RNA sequencing (RNA-seq) datasets, uniquely obtained from human and murine embryonic hearts. We found that while STRA6 mRNA was much less frequently expressed in murine embryonic heart cells derived from the Mesp1+ lineage tracing mice (Mesp1Cre/+; Rosa26tdTomato), it was expressed predominantly in the OFT region-specific heart progenitors in human developing hearts. Next, we revealed that STRA6-knockout human embryonic stem cells (hESCs) could differentiate into cardiomyocytes similarly to wild-type hESCs, but could not differentiate properly into mesodermal nor neural crest cell-derived smooth muscle cells (SMCs) in vitro. This is supported by the population RNA-seq data showing down-regulation of the SMC-related genes in the STRA6-knockout hESC-derived cells. Further, through machinery assays, we identified the previously unrecognized interaction between RA nuclear receptors RARα/RXRα and TBX1, an OFT-specific cardiogenic transcription factor, which would likely act downstream to STRA6-mediated RA signalling in human cardiogenesis.
Conclusion: Our study highlights the critical role of human-specific STRA6 progenitors for proper induction of vascular SMCs that is essential for normal OFT formation. Thus, these results shed light on novel and human-specific CHD mechanisms, driven by STRA6 mutations.
Keywords: Cardiac outflow tract; Congenital heart disease; Heart development; Matthew-Wood syndrome; Retinoic acid; Single-cell RNA-seq; Smooth muscle cell.
© The Author(s) 2023. Published by Oxford University Press on behalf of the European Society of Cardiology.
Conflict of interest statement
Conflict of interest: The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Vincent SD, Buckingham ME. How to make a heart: the origin and regulation of cardiac progenitor cells. Curr Top Dev Biol 2010;90:1–41. - PubMed
-
- Sakabe M, Kokubo H, Nakajima Y, Saga Y. Ectopic retinoic acid signaling affects outflow tract cushion development through suppression of the myocardial Tbx2-Tgfβ2 pathway. Development 2012;139:385–395. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
