

GOT1 regulates CD8+ effector and memory T cell generation
- PMID: 36640309
- PMCID: PMC9943022
- DOI: 10.1016/j.celrep.2022.111987
GOT1 regulates CD8+ effector and memory T cell generation
Abstract
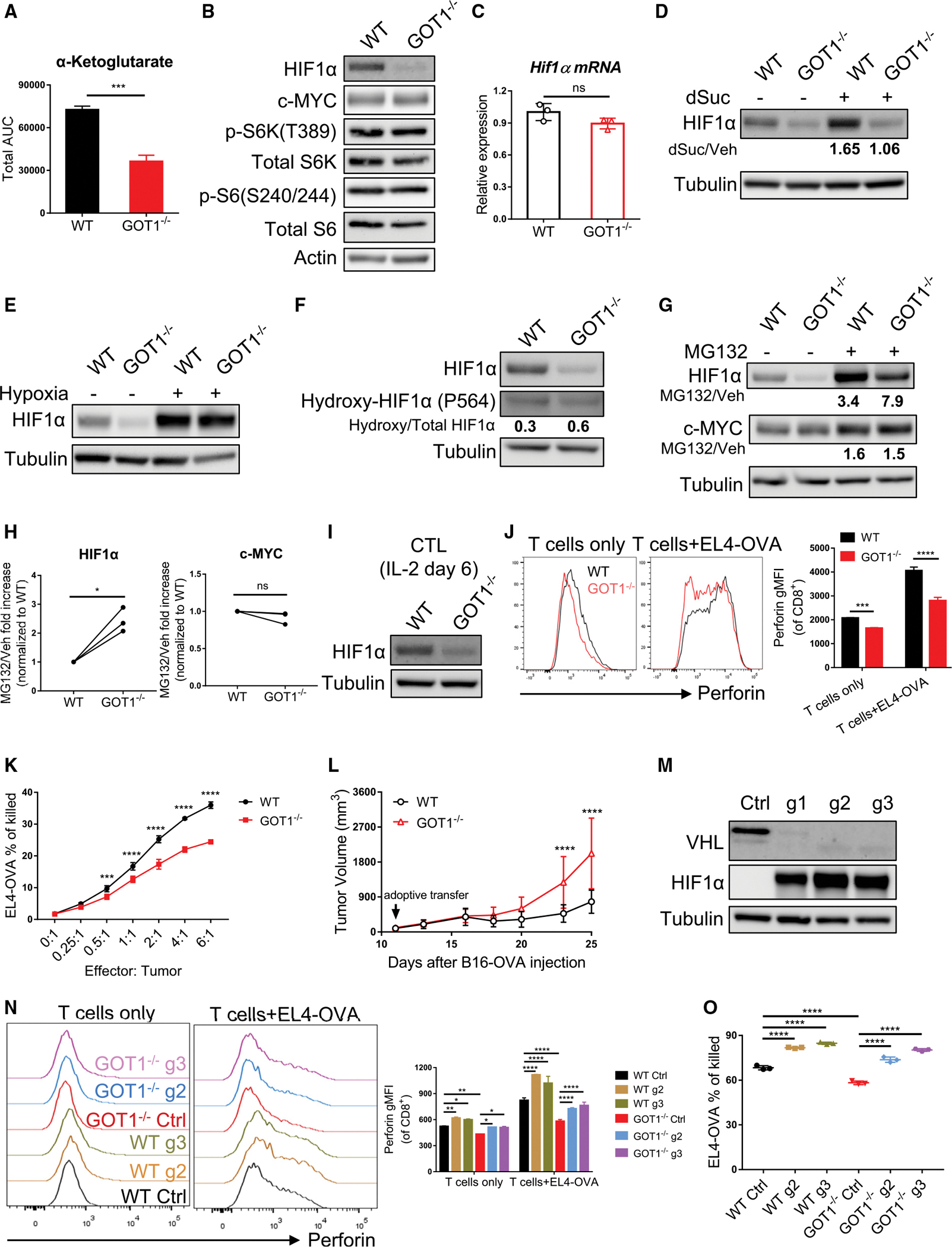
T cell activation, proliferation, function, and differentiation are tightly linked to proper metabolic reprogramming and regulation. By using [U-13C]glucose tracing, we reveal a critical role for GOT1 in promoting CD8+ T cell effector differentiation and function. Mechanistically, GOT1 enhances proliferation by maintaining intracellular redox balance and serine-mediated purine nucleotide biosynthesis. Further, GOT1 promotes the glycolytic programming and cytotoxic function of cytotoxic T lymphocytes via posttranslational regulation of HIF protein, potentially by regulating the levels of α-ketoglutarate. Conversely, genetic deletion of GOT1 promotes the generation of memory CD8+ T cells.
Keywords: CP: Metabolism; GOT1; HIF; NADH/NAD; effector and memory CD8(+) T cell; glucose; glutamate; serine.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.D.P. is a cofounder and equity holder of Dracen Pharmaceuticals. C.H.P. and J.D.P. are current employees of Calico LLC.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
