

Chemokine CXCL4 interactions with extracellular matrix proteoglycans mediate widespread immune cell recruitment independent of chemokine receptors
- PMID: 36640356
- PMCID: PMC11064100
- DOI: 10.1016/j.celrep.2022.111930
Chemokine CXCL4 interactions with extracellular matrix proteoglycans mediate widespread immune cell recruitment independent of chemokine receptors
Abstract
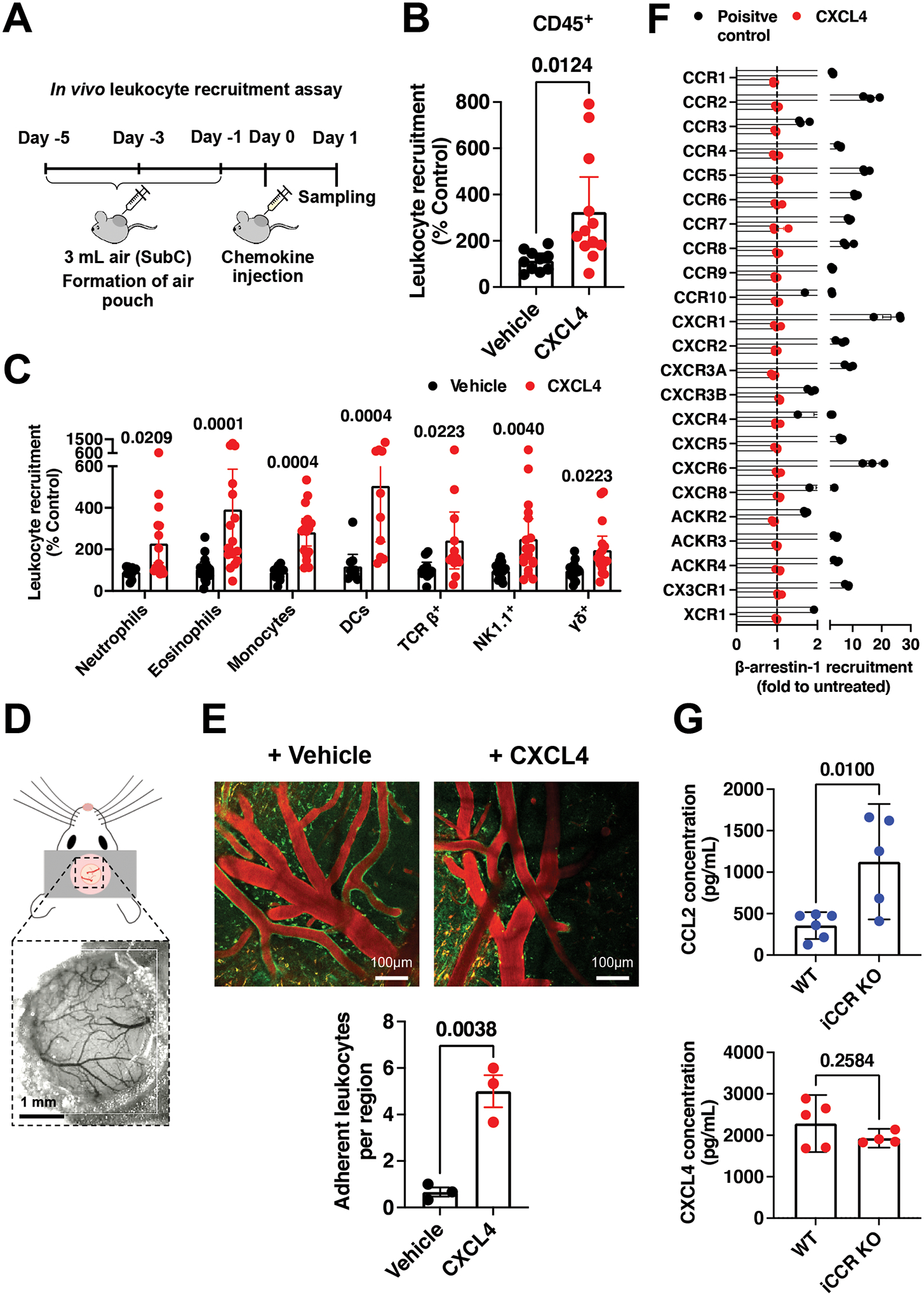
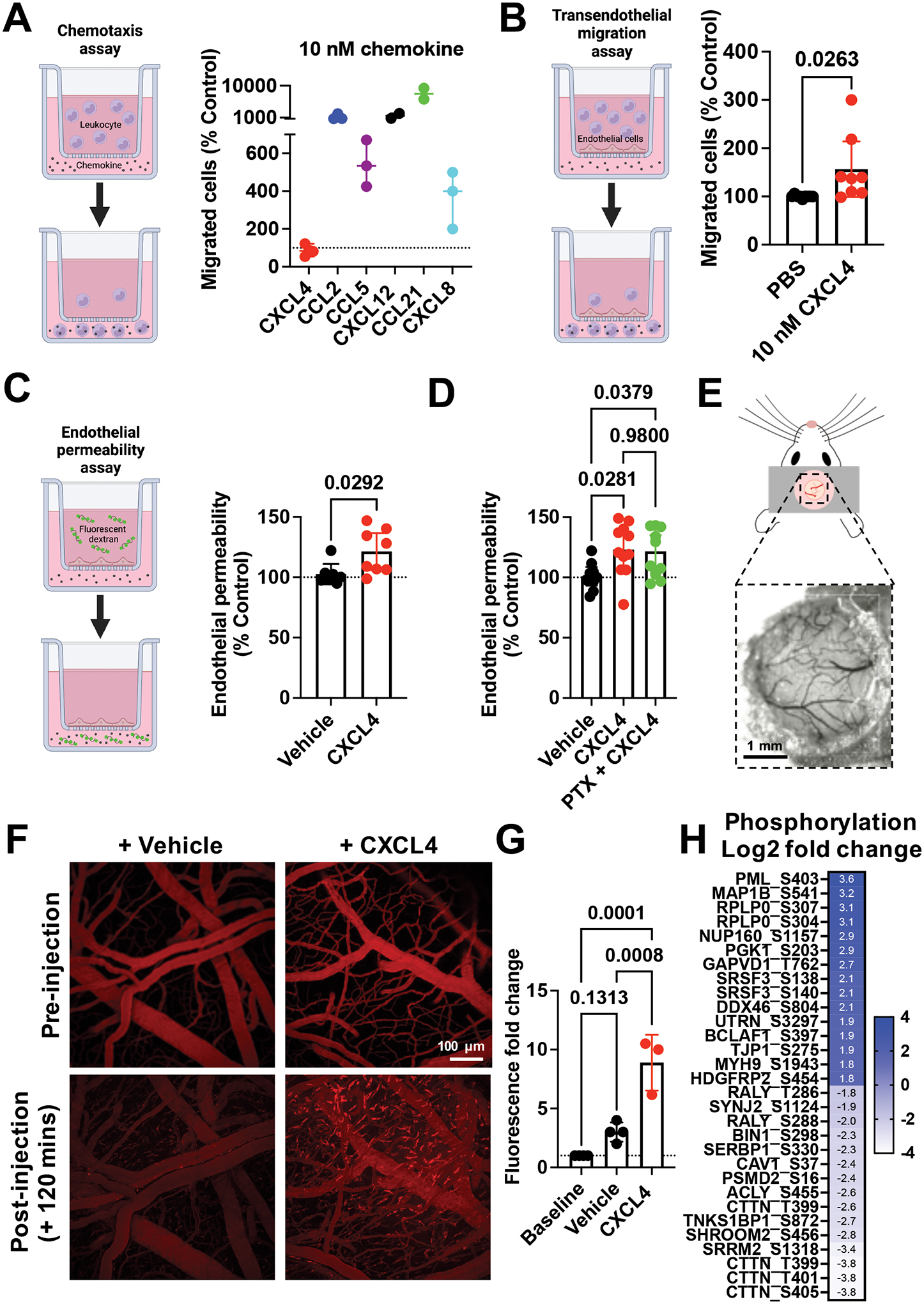
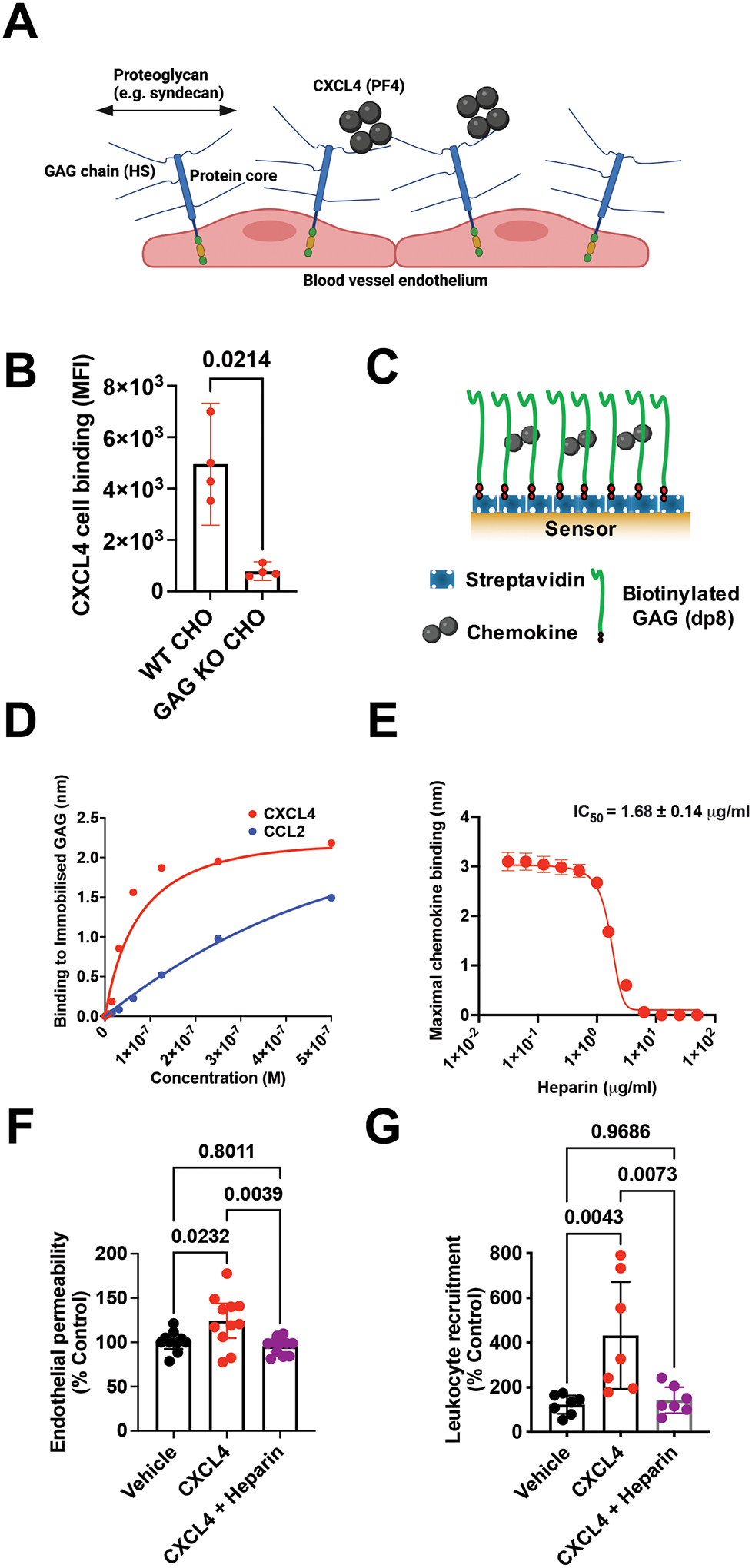
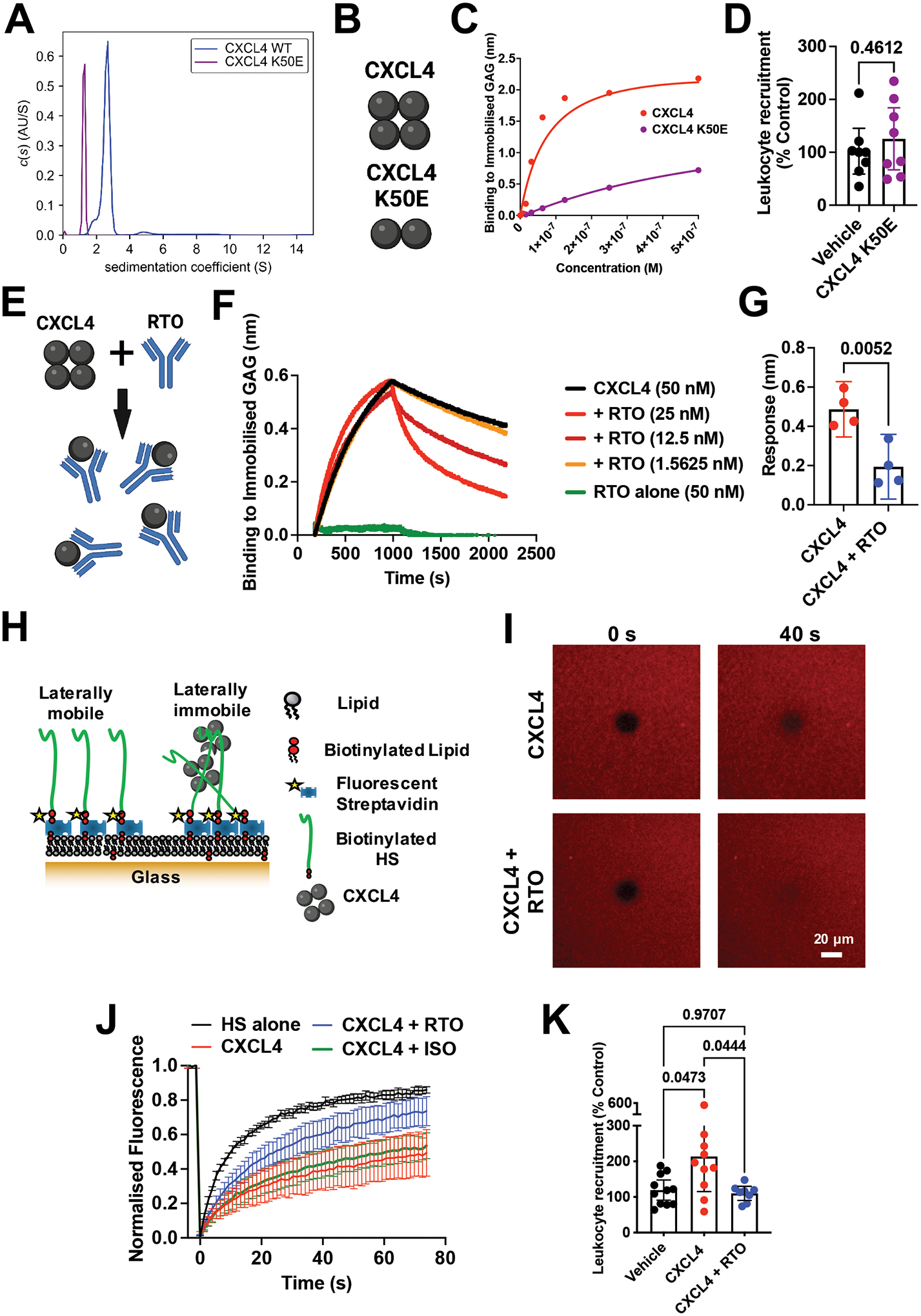
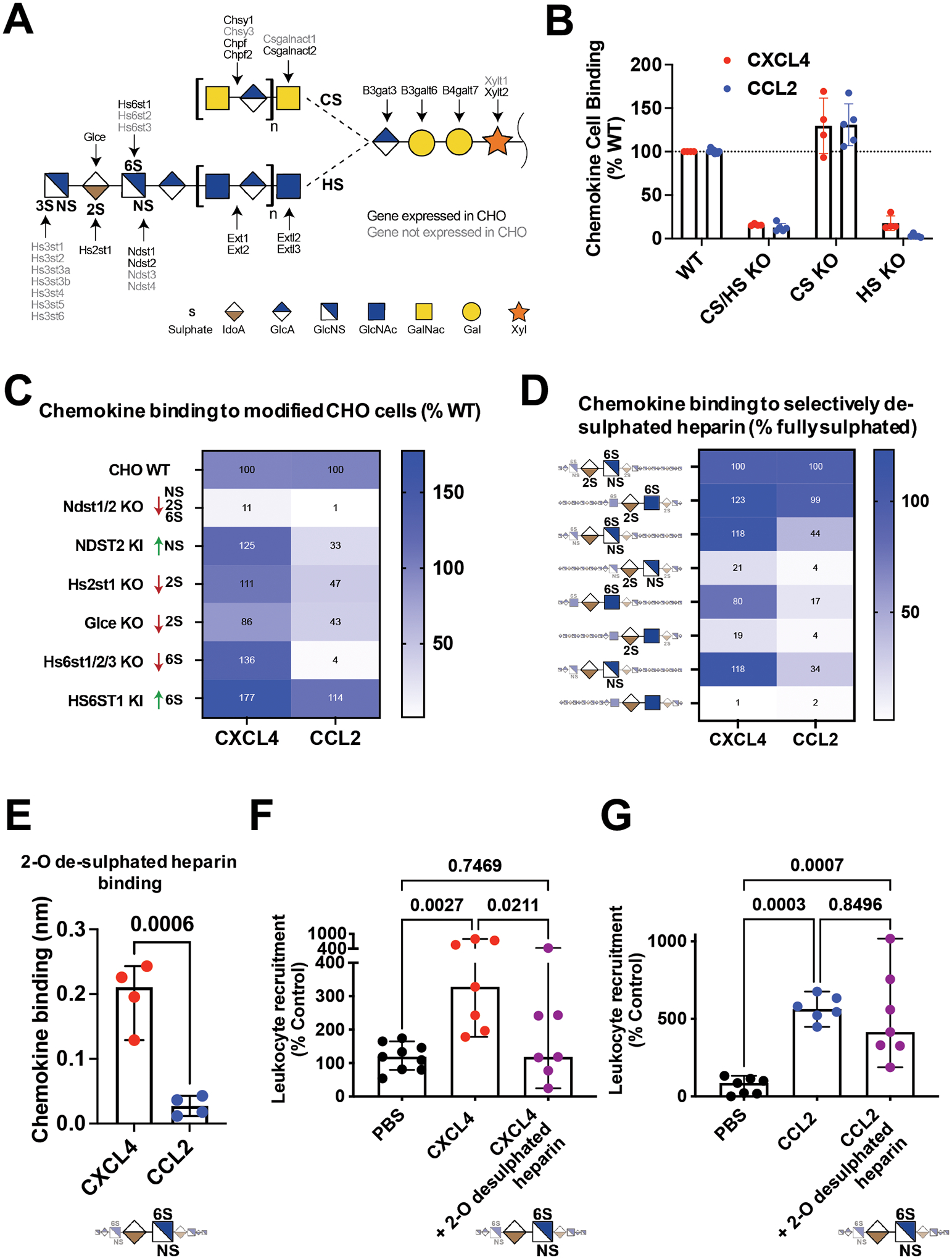
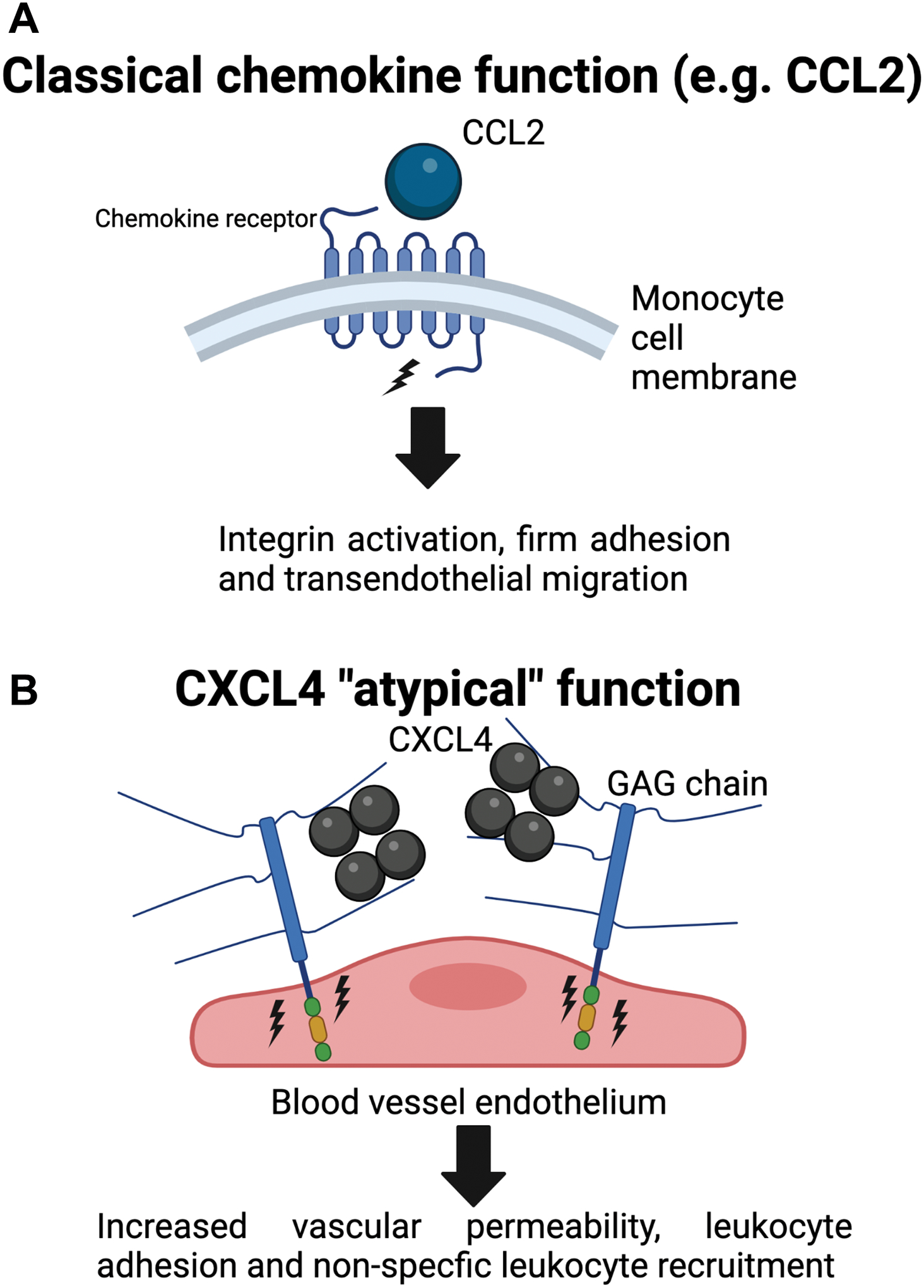
Leukocyte recruitment from the vasculature into tissues is a crucial component of the immune system but is also key to inflammatory disease. Chemokines are central to this process but have yet to be therapeutically targeted during inflammation due to a lack of mechanistic understanding. Specifically, CXCL4 (Platelet Factor 4, PF4) has no established receptor that explains its function. Here, we use biophysical, in vitro, and in vivo techniques to determine the mechanism underlying CXCL4-mediated leukocyte recruitment. We demonstrate that CXCL4 binds to glycosaminoglycan (GAG) sugars on proteoglycans within the endothelial extracellular matrix, resulting in increased adhesion of leukocytes to the vasculature, increased vascular permeability, and non-specific recruitment of a range of leukocytes. Furthermore, GAG sulfation confers selectivity onto chemokine localization. These findings present mechanistic insights into chemokine biology and provide future therapeutic targets.
Keywords: CP: Immunology; CXCL4; PF4; chemokine; extracellular matrix; glycosaminoglycan; leukocyte; proteoglycan; recruitment; trafficking.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Bachelerie F, Ben-Baruch A, Burkhardt AM, Combadière C, Farber JM, Graham GJ, Horuk R, Sparre-Ulrich AH, Locati M, Luster AD, et al. (2014). International Union of Basic and Clinical Pharmacology. [corrected]. LXXXIX. Update on the extended family of chemokine receptors and introducing a new nomenclature for atypical chemokine receptors. Pharmacological Reviews 66, 1–79. 10.1124/pr.113.007724. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
