

Use of 2,6-diaminopurine as a potent suppressor of UGA premature stop codons in cystic fibrosis
- PMID: 36641622
- PMCID: PMC10124085
- DOI: 10.1016/j.ymthe.2023.01.014
Use of 2,6-diaminopurine as a potent suppressor of UGA premature stop codons in cystic fibrosis
Abstract
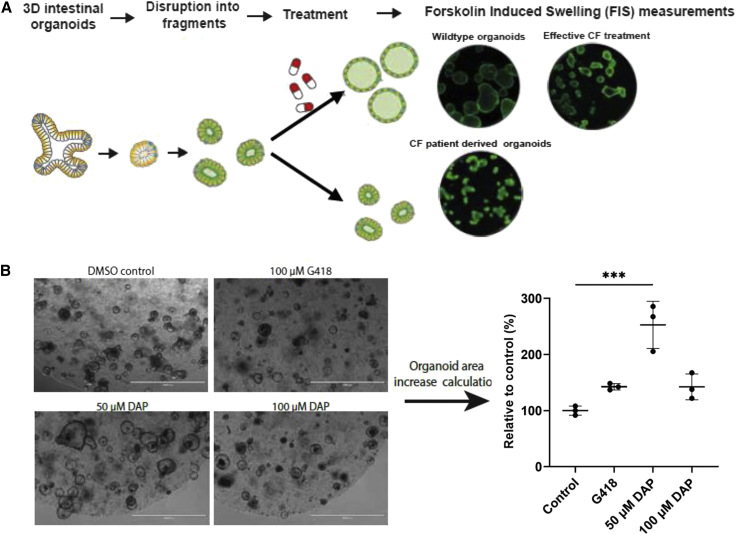
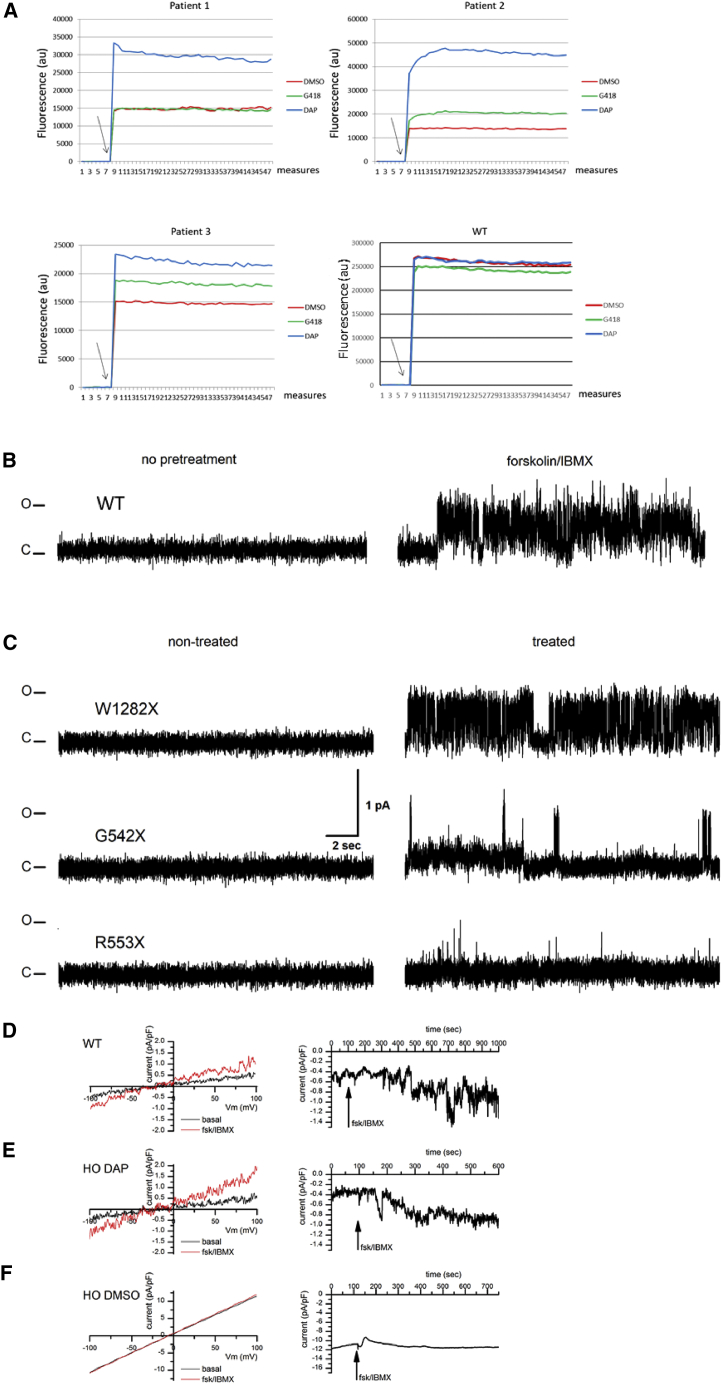
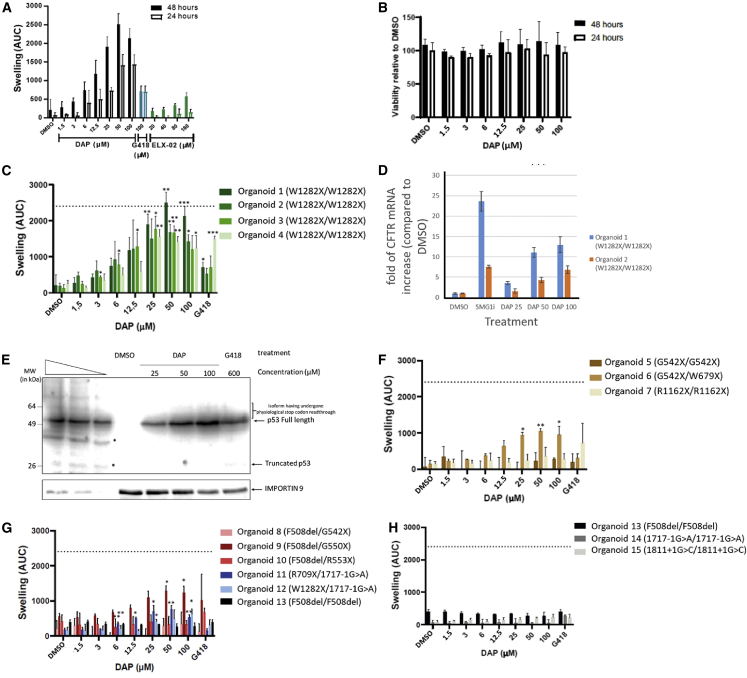
Nonsense mutations are responsible for around 10% of cases of genetic diseases, including cystic fibrosis. 2,6-diaminopurine (DAP) has recently been shown to promote efficient readthrough of UGA premature stop codons. In this study, we show that DAP can correct a nonsense mutation in the Cftr gene in vivo in a new CF mouse model, in utero, and through breastfeeding, thanks, notably, to adequate pharmacokinetic properties. DAP turns out to be very stable in plasma and is distributed throughout the body. The ability of DAP to correct various endogenous UGA nonsense mutations in the CFTR gene and to restore its function in mice, in organoids derived from murine or patient cells, and in cells from patients with cystic fibrosis reveals the potential of such readthrough-stimulating molecules in developing a therapeutic approach. The fact that correction by DAP of certain nonsense mutations reaches a clinically relevant level, as judged from previous studies, makes the use of this compound all the more attractive.
Keywords: 2,6-diaminopurine; cystic fibrosis; mouse model; nonsense mutation; readthrough molecule.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.B. is inventor on a patent related to the FIS assay and received financial royalties from 2017 onward. S.A., C. Bailly, S.R., and F.L. are inventors on a patent demonstrating that DAP is a readthrough molecule useful for the treatment of genetic diseases related to nonsense mutations.
Figures
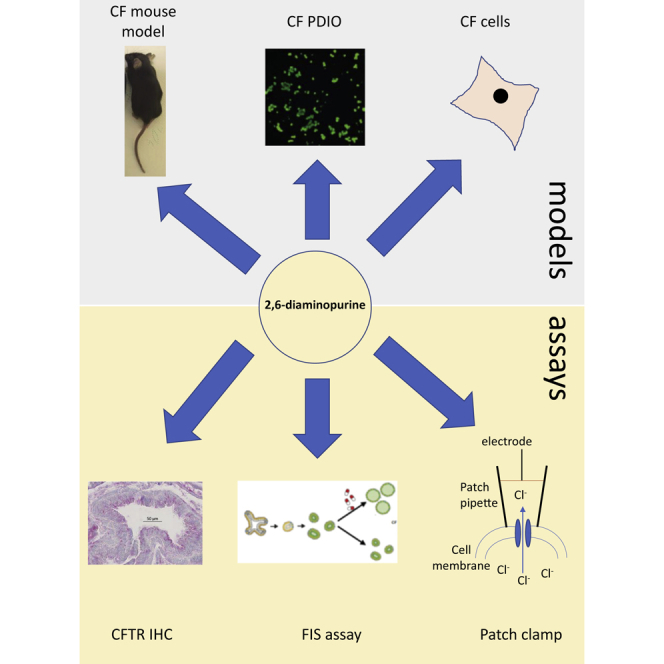
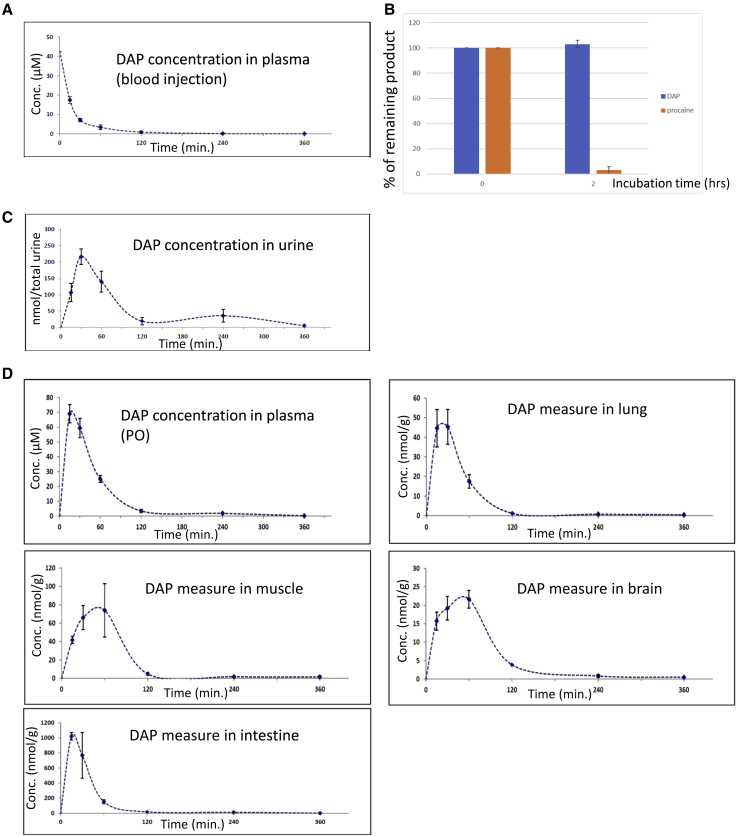
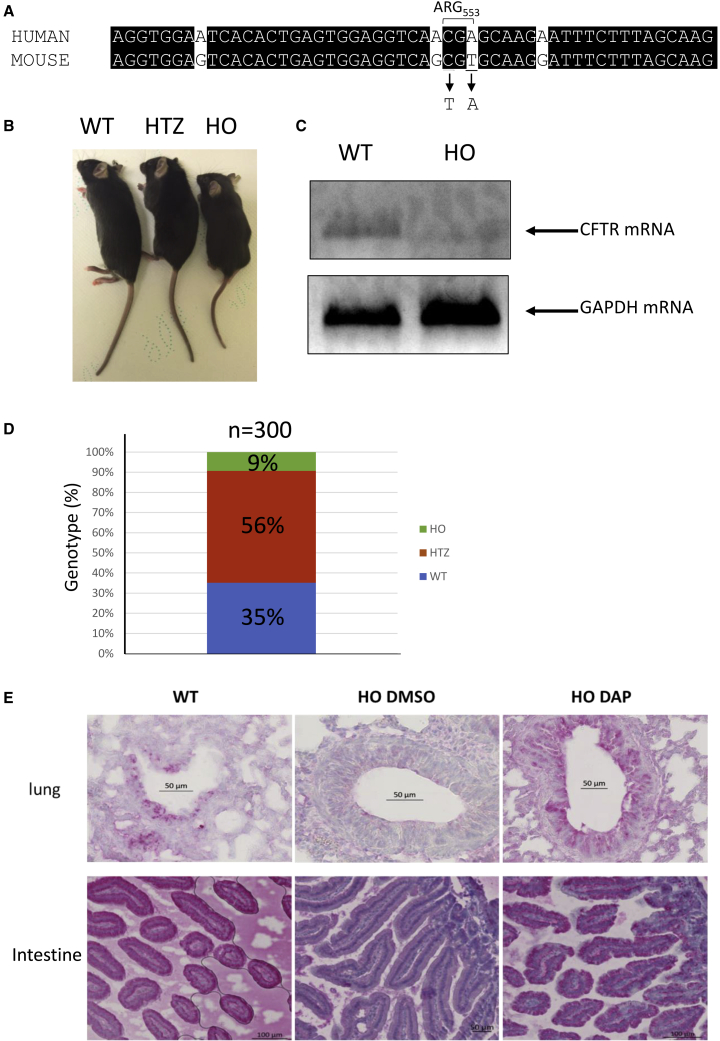
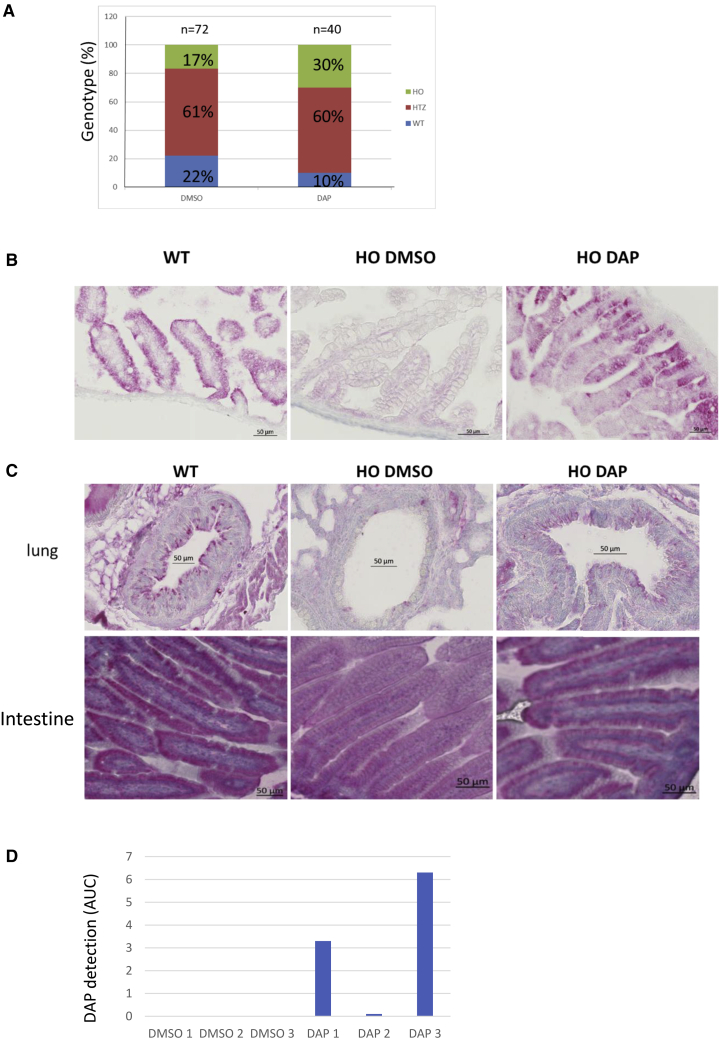
Comment in
-
The power of 2,6-diaminopurine in correcting UGA nonsense codons in CFTR mRNA.Mol Ther. 2023 Apr 5;31(4):921-922. doi: 10.1016/j.ymthe.2023.03.009. Epub 2023 Mar 21. Mol Ther. 2023. PMID: 36944336 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
