

MCT1-dependent energetic failure and neuroinflammation underlie optic nerve degeneration in Wolfram syndrome mice
- PMID: 36645345
- PMCID: PMC9891717
- DOI: 10.7554/eLife.81779
MCT1-dependent energetic failure and neuroinflammation underlie optic nerve degeneration in Wolfram syndrome mice
Abstract
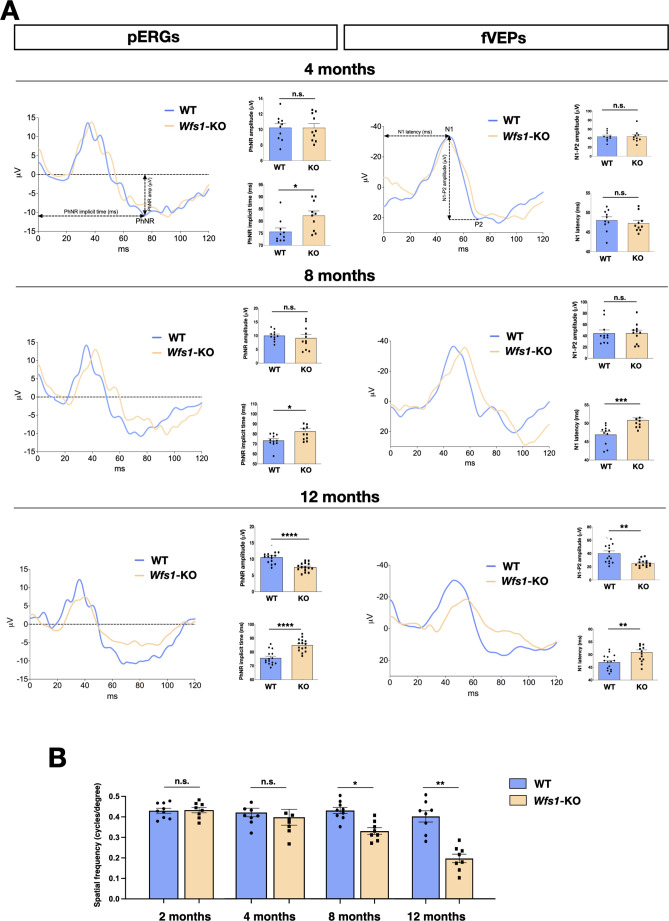
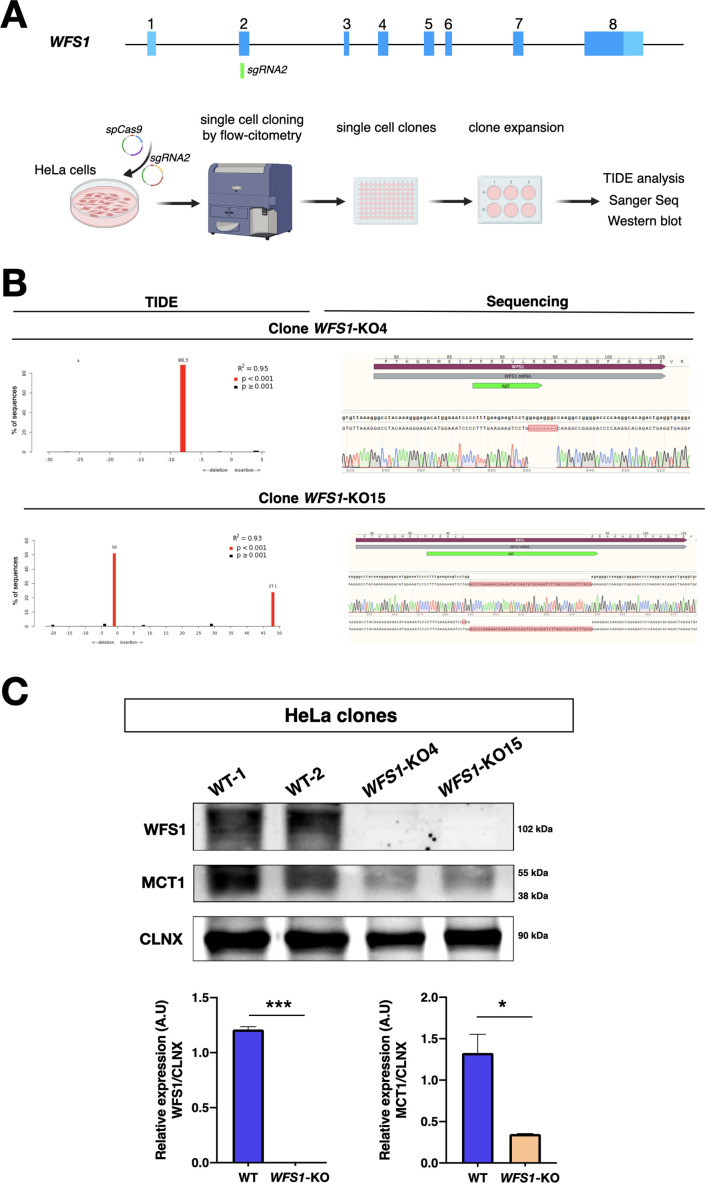
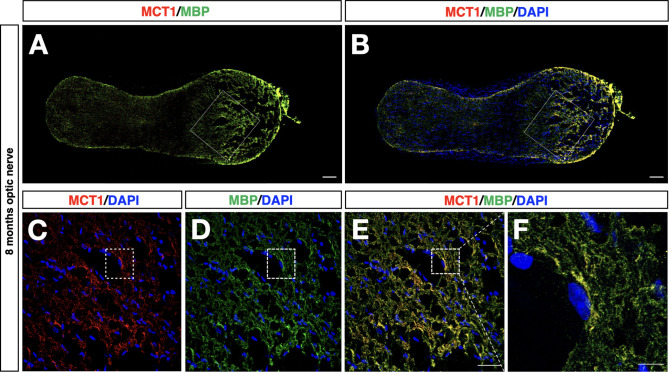
Wolfram syndrome 1 (WS1) is a rare genetic disorder caused by mutations in the WFS1 gene leading to a wide spectrum of clinical dysfunctions, among which blindness, diabetes, and neurological deficits are the most prominent. WFS1 encodes for the endoplasmic reticulum (ER) resident transmembrane protein wolframin with multiple functions in ER processes. However, the WFS1-dependent etiopathology in retinal cells is unknown. Herein, we showed that Wfs1 mutant mice developed early retinal electrophysiological impairments followed by marked visual loss. Interestingly, axons and myelin disruption in the optic nerve preceded the degeneration of the retinal ganglion cell bodies in the retina. Transcriptomics at pre-degenerative stage revealed the STAT3-dependent activation of proinflammatory glial markers with reduction of the homeostatic and pro-survival factors glutamine synthetase and BDNF. Furthermore, label-free comparative proteomics identified a significant reduction of the monocarboxylate transport isoform 1 (MCT1) and its partner basigin that are highly enriched on retinal glia and myelin-forming oligodendrocytes in optic nerve together with wolframin. Loss of MCT1 caused a failure in lactate transfer from glial to neuronal cell bodies and axons leading to a chronic hypometabolic state. Thus, this bioenergetic impairment is occurring concurrently both within the axonal regions and cell bodies of the retinal ganglion cells, selectively endangering their survival while impacting less on other retinal cells. This metabolic dysfunction occurs months before the frank RGC degeneration suggesting an extended time-window for intervening with new therapeutic strategies focused on boosting retinal and optic nerve bioenergetics in WS1.
Keywords: astrocytes; developmental biology; eye; glial cell; mouse; retinal development.
© 2023, Rossi et al.
Conflict of interest statement
GR, GO, NV, VC, AI, DD, EB, LB, SG, ML, SM, LL, PM, VB No competing interests declared
Figures








References
-
- Angebault C, Fauconnier J, Patergnani S, Rieusset J, Danese A, Affortit CA, Jagodzinska J, Mégy C, Quiles M, Cazevieille C, Korchagina J, Bonnet-Wersinger D, Milea D, Hamel C, Pinton P, Thiry M, Lacampagne A, Delprat B, Delettre C. ER-mitochondria cross-talk is regulated by the ca2+ sensor NCS1 and is impaired in wolfram syndrome. Science Signaling. 2018;11:eaaq1380. doi: 10.1126/scisignal.aaq1380. - DOI - PubMed
-
- Barboni P, Amore G, Cascavilla ML, Battista M, Frontino G, Romagnoli M, Caporali L, Baldoli C, Gramegna LL, Sessagesimi E, Bonfanti R, Romagnoli A, Scotti R, Brambati M, Carbonelli M, Starace V, Fiorini C, Panebianco R, Parisi V, Tonon C, Bandello F, Carelli V, La Morgia C. The pattern of retinal ganglion cell loss in Wolfram syndrome is distinct from mitochondrial optic neuropathies. American Journal of Ophthalmology. 2022;241:206–216. doi: 10.1016/j.ajo.2022.03.019. - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
