

Molecular fate-mapping of serum antibody responses to repeat immunization
- PMID: 36646114
- PMCID: PMC10023323
- DOI: 10.1038/s41586-023-05715-3
Molecular fate-mapping of serum antibody responses to repeat immunization
Abstract
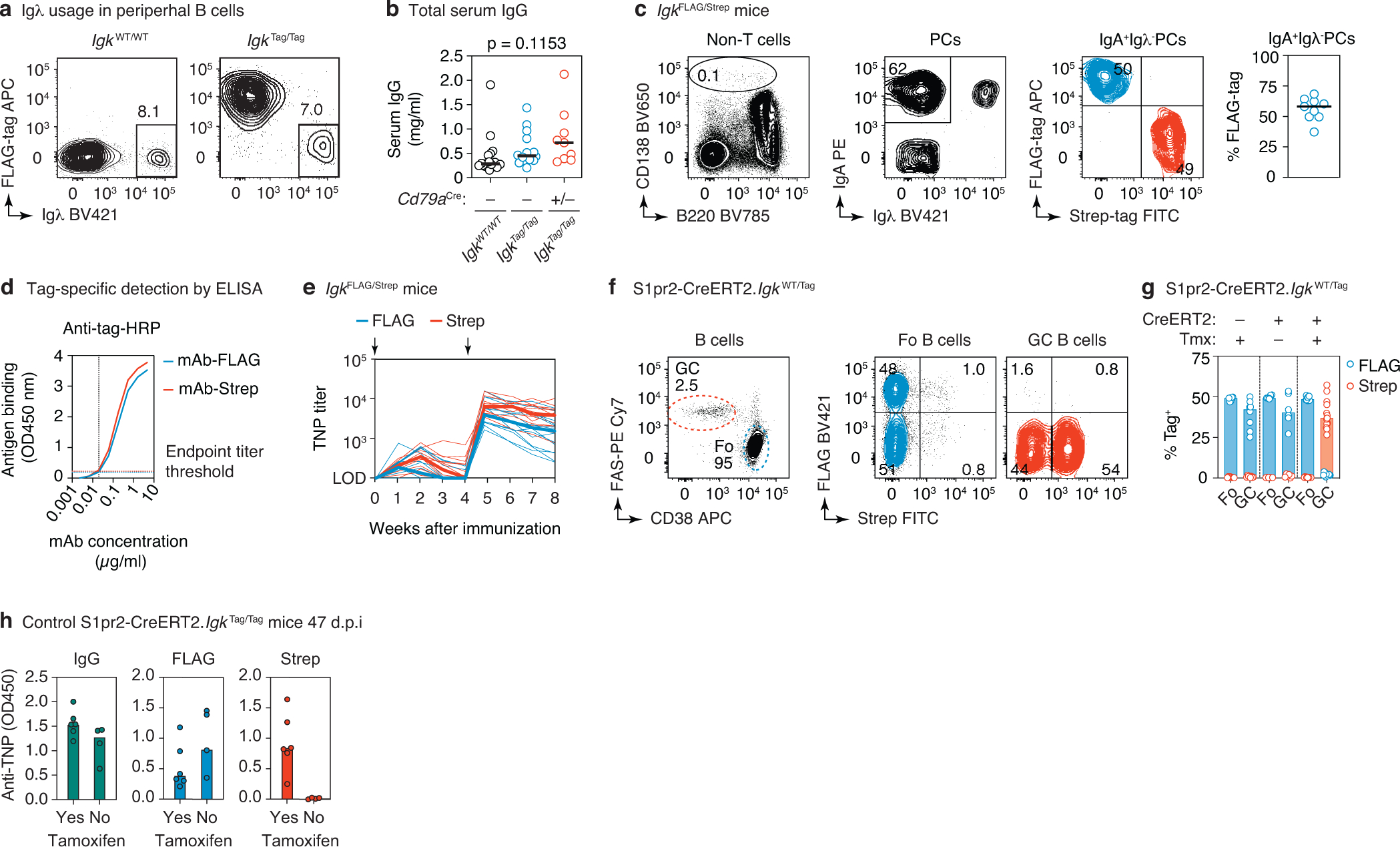
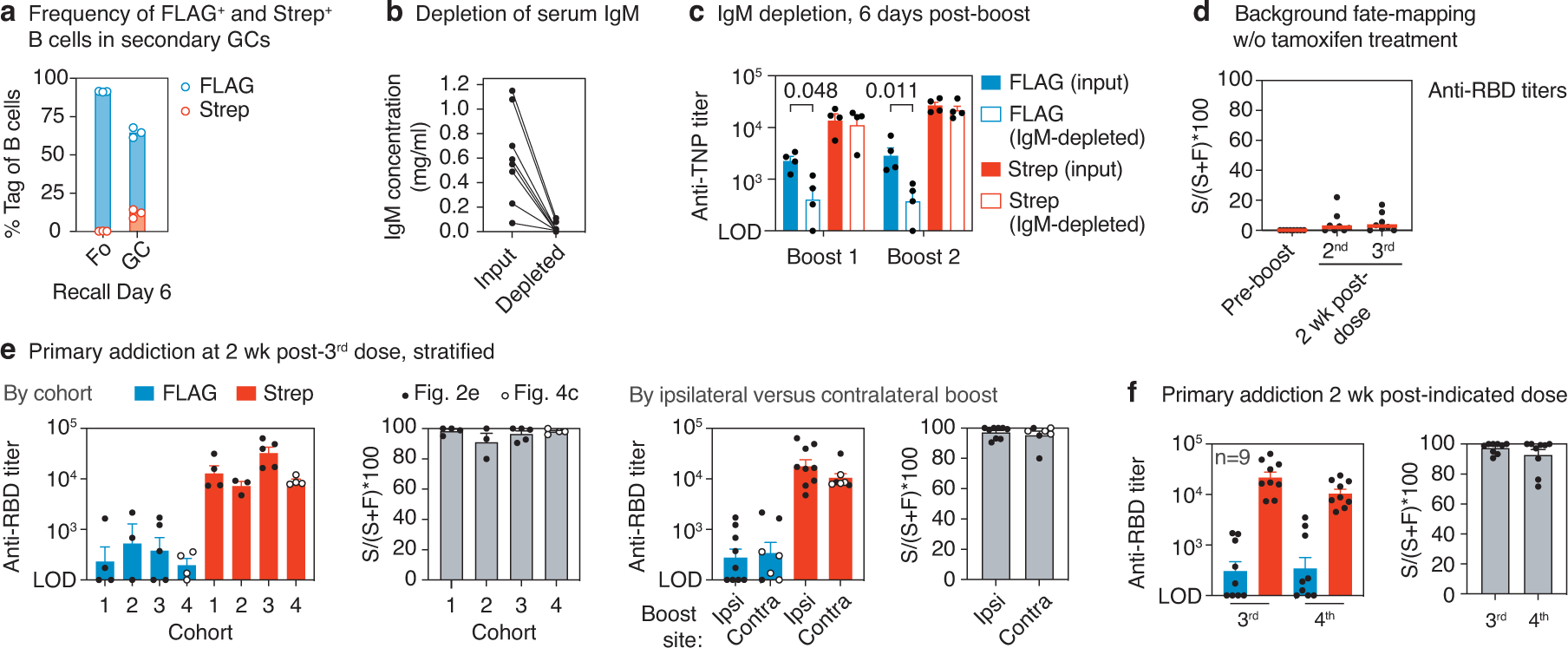
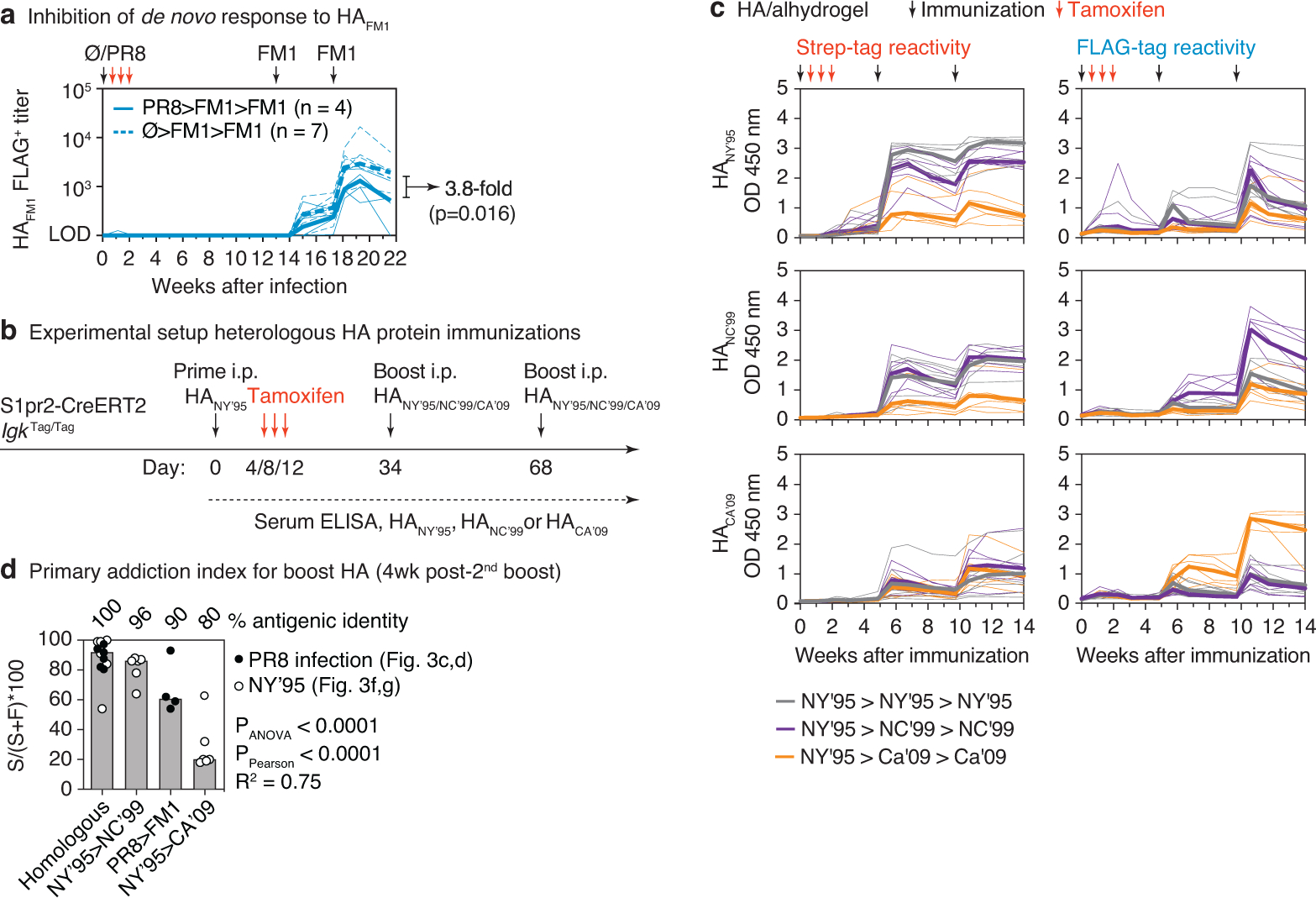
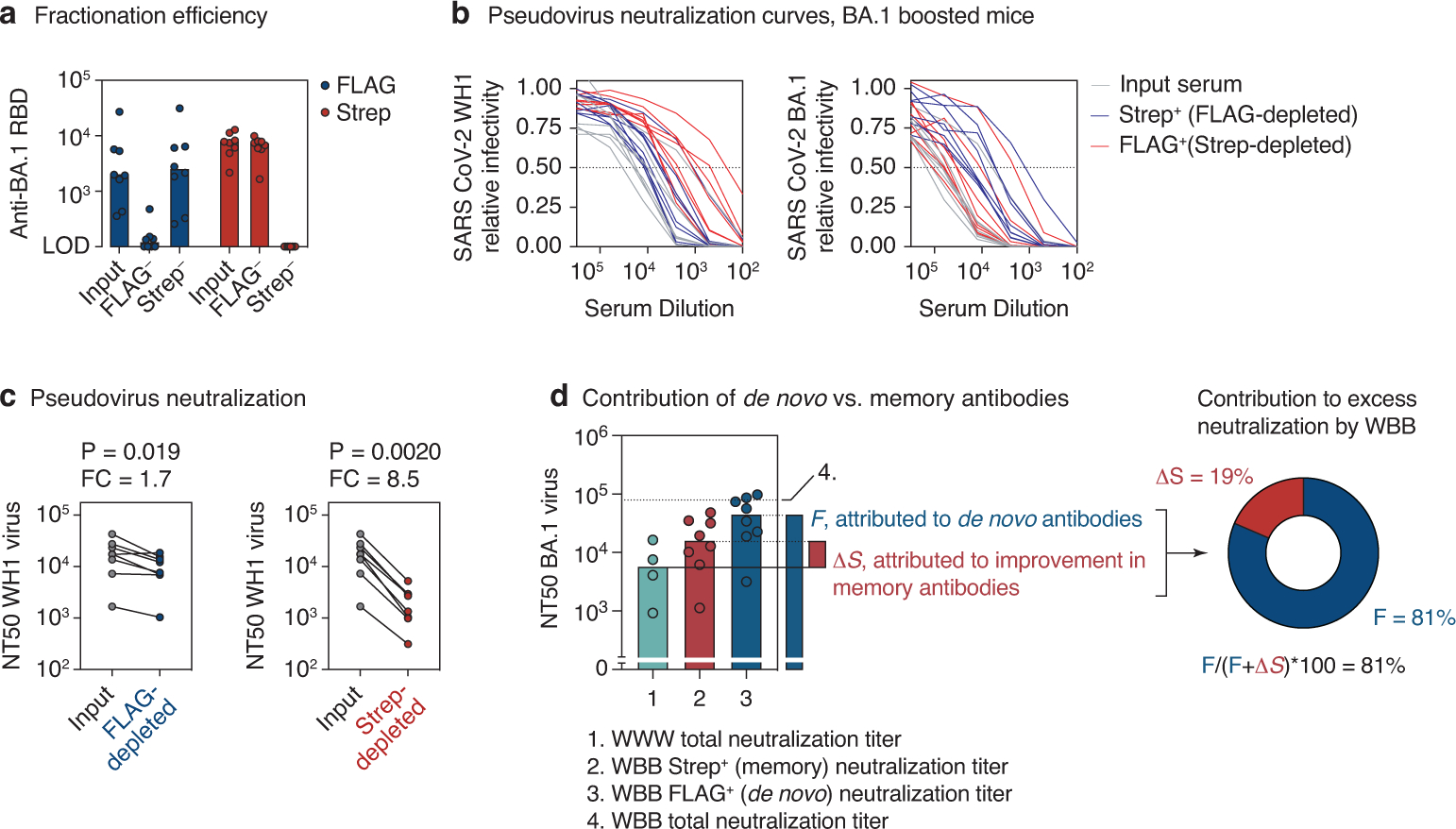
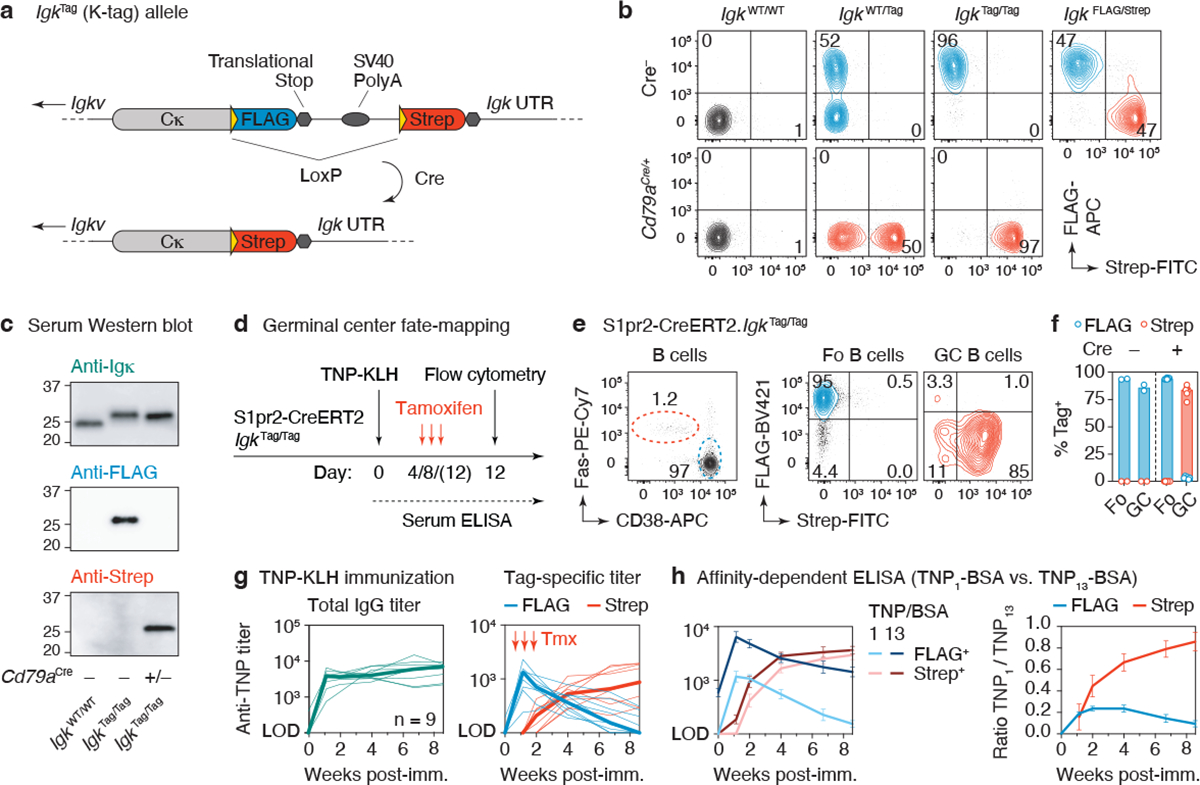
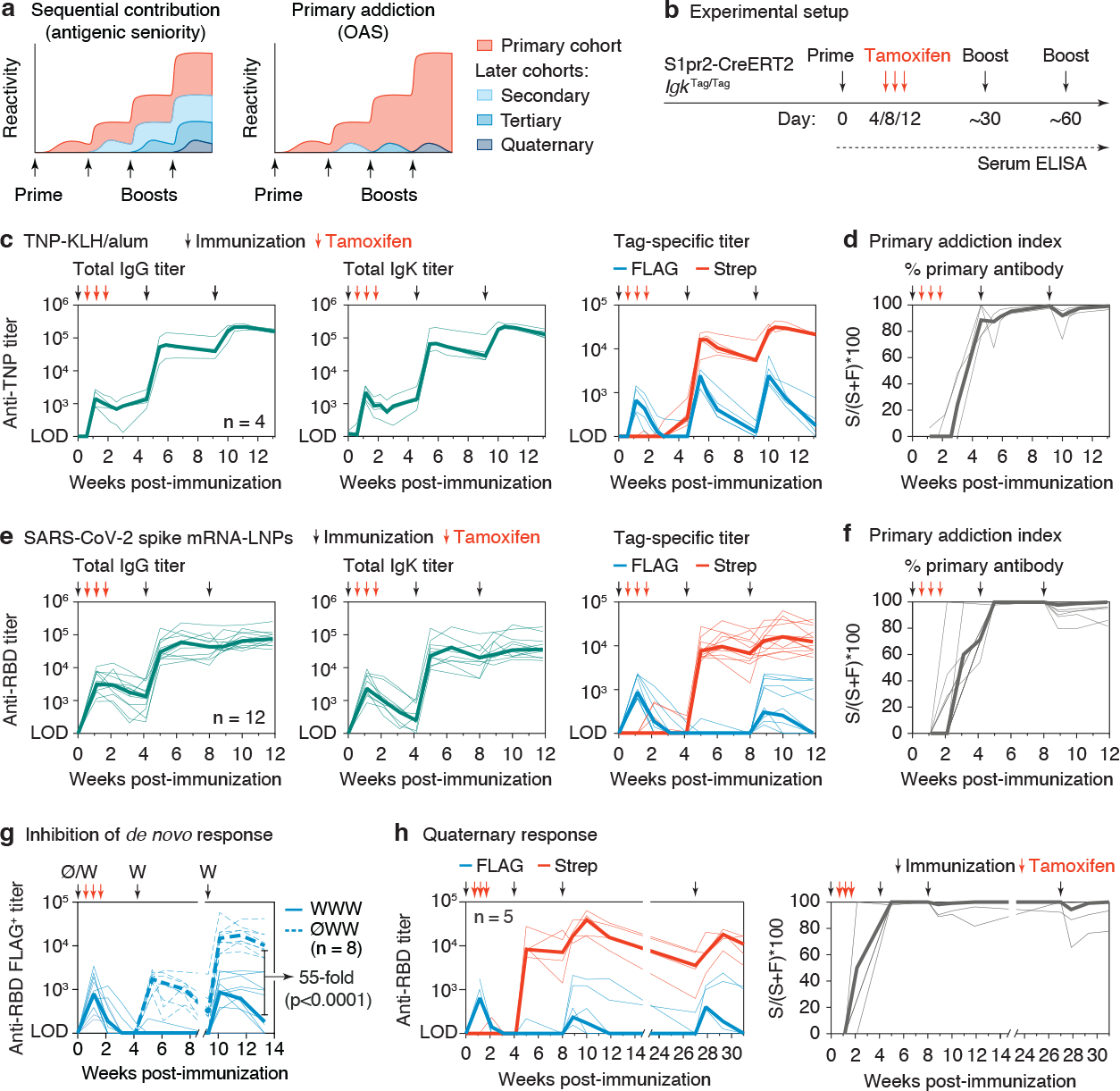
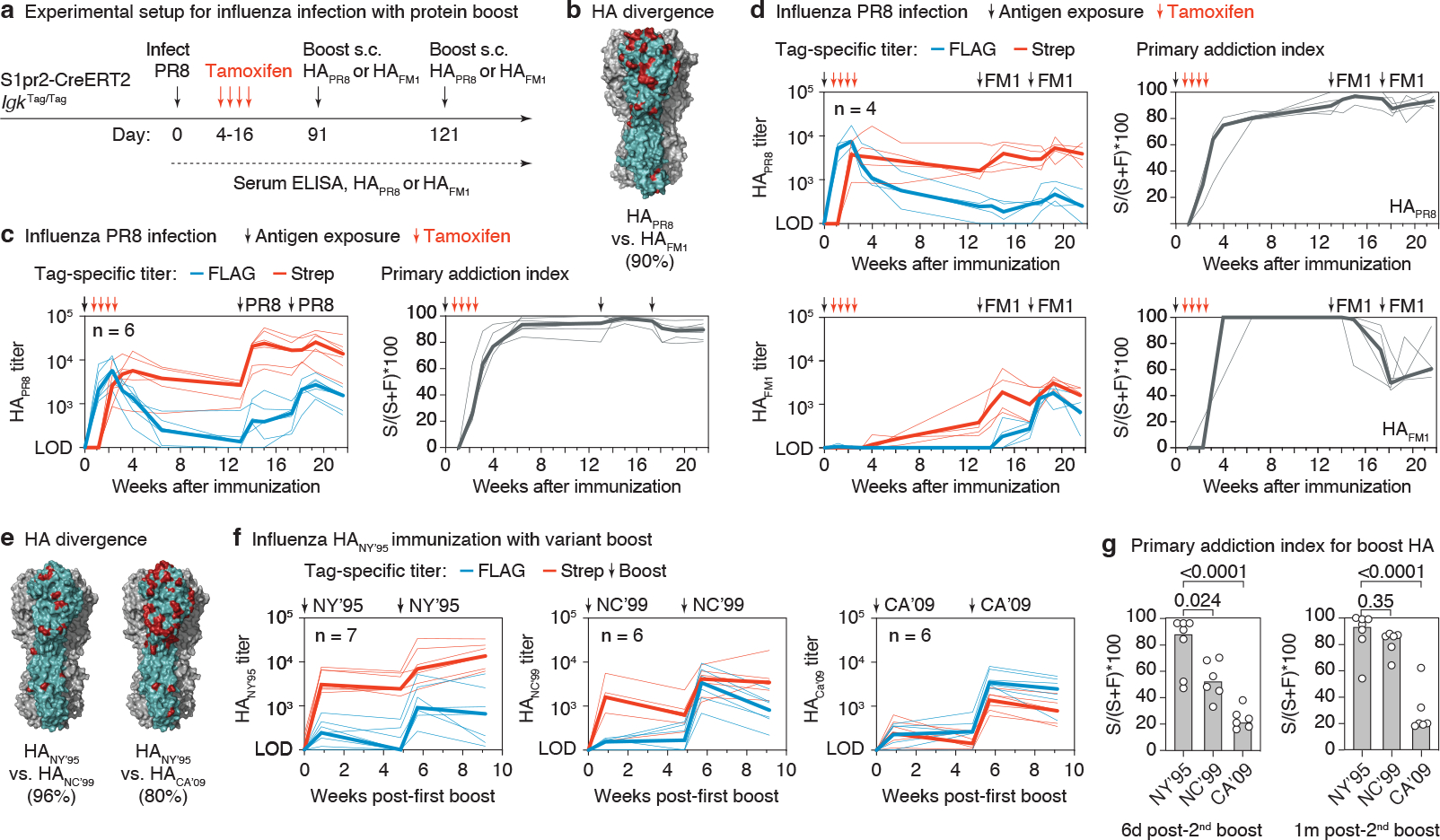
The protective efficacy of serum antibodies results from the interplay of antigen-specific B cell clones of different affinities and specificities. These cellular dynamics underlie serum-level phenomena such as original antigenic sin (OAS)-a proposed propensity of the immune system to rely repeatedly on the first cohort of B cells engaged by an antigenic stimulus when encountering related antigens, in detriment to the induction of de novo responses1-5. OAS-type suppression of new, variant-specific antibodies may pose a barrier to vaccination against rapidly evolving viruses such as influenza and SARS-CoV-26,7. Precise measurement of OAS-type suppression is challenging because cellular and temporal origins cannot readily be ascribed to antibodies in circulation; its effect on subsequent antibody responses therefore remains unclear5,8. Here we introduce a molecular fate-mapping approach with which serum antibodies derived from specific cohorts of B cells can be differentially detected. We show that serum responses to sequential homologous boosting derive overwhelmingly from primary cohort B cells, while later induction of new antibody responses from naive B cells is strongly suppressed. Such 'primary addiction' decreases sharply as a function of antigenic distance, allowing reimmunization with divergent viral glycoproteins to produce de novo antibody responses targeting epitopes that are absent from the priming variant. Our findings have implications for the understanding of OAS and for the design and testing of vaccines against evolving pathogens.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
N.P. is named on a patent describing the use of nucleoside-modified mRNA in lipid nanoparticles as a vaccine platform. He has disclosed those interests fully to the University of Pennsylvania and has an approved plan in place for managing any potential conflicts arising from the licensing of that patent. Paulo J.C. Lin and Ying K. Tam are employees of Acuitas Therapeutics, a company involved in the development of mRNA-LNP therapeutics. Ying K. Tam is named on patents that describe lipid nanoparticles for the delivery of nucleic acid therapeutics, including mRNA, and the use of modified mRNA in lipid nanoparticles as a vaccine platform. P.D.B. has done consulting work in the area of COVID vaccines for Pfizer Inc.. J.D.B. consults or has recently consulted for Apriori Bio, Oncorus, Merck, and Moderna on topics related to viruses, vaccines, and viral evolution. J.D.B, T.N.S., and A.J.G. are inventors on Fred Hutch licensed patents related to viral deep mutational scanning. G.D.V. and J.D.B. are advisors for the Vaccine Company, Inc..
Figures




Comment in
-
Plasmablasts from the past: Nostalgic B cells can't let go.Sci Immunol. 2023 Mar 10;8(81):eadh3115. doi: 10.1126/sciimmunol.adh3115. Epub 2023 Mar 3. Sci Immunol. 2023. PMID: 36867677
-
Total recall? Understanding the effect of antigenic distance on original antigenic sin.Immunol Cell Biol. 2023 May;101(5):377-380. doi: 10.1111/imcb.12638. Epub 2023 Mar 27. Immunol Cell Biol. 2023. PMID: 36880370
-
Fate-mapping antibodies to study sinful immune dynamics.Nat Immunol. 2023 Apr;24(4):570-572. doi: 10.1038/s41590-023-01467-4. Nat Immunol. 2023. PMID: 36959294 No abstract available.
-
Opposing effects of pre-existing antibody and memory T cell help on the dynamics of recall germinal centers.Immunity. 2024 Jul 9;57(7):1618-1628.e4. doi: 10.1016/j.immuni.2024.05.009. Epub 2024 Jun 4. Immunity. 2024. PMID: 38838672 Free PMC article.
References
MAIN TEXT REFERENCES
-
- Francis T On the Doctrine of Original Antigenic Sin. Proceedings of the American Philosophical Society 104, 572–578 (1960).
ADDITIONAL REFERENCES
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
