

Epigenetic and post-translational modifications in autophagy: biological functions and therapeutic targets
- PMID: 36646695
- PMCID: PMC9842768
- DOI: 10.1038/s41392-022-01300-8
Epigenetic and post-translational modifications in autophagy: biological functions and therapeutic targets
Abstract
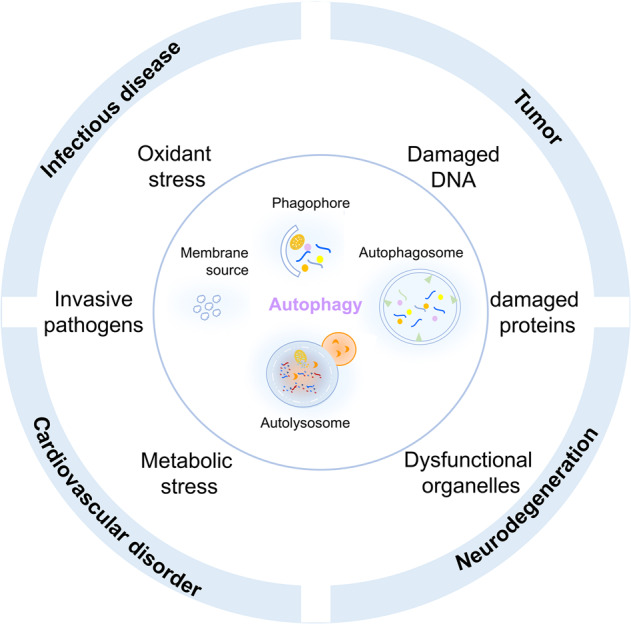
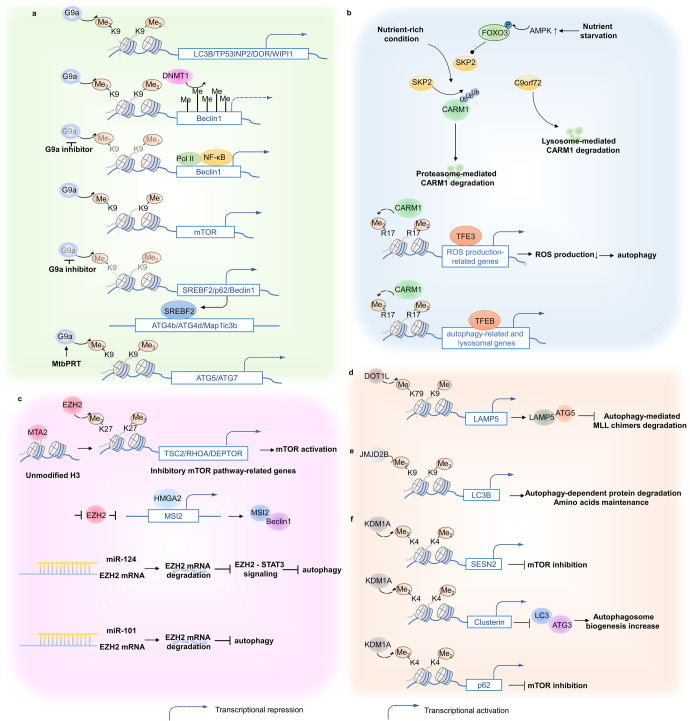
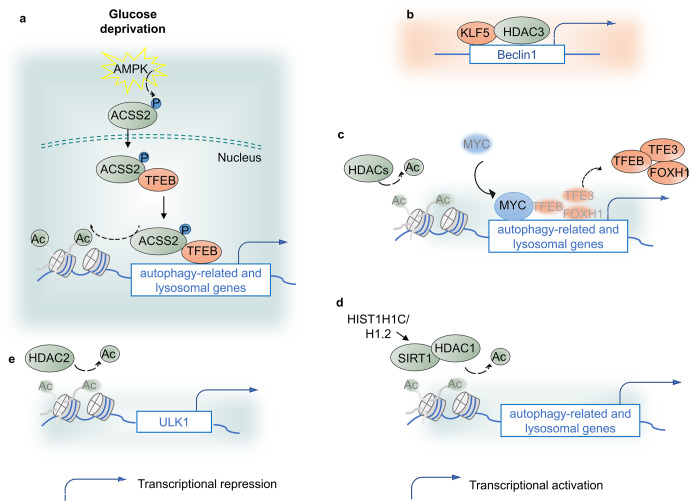
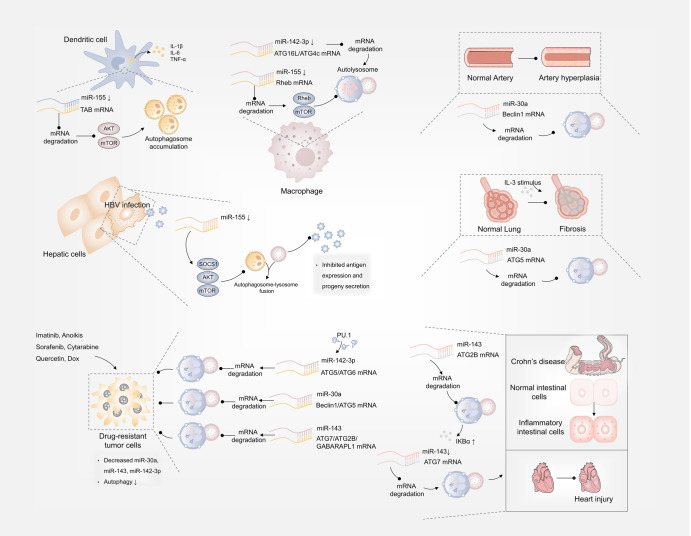
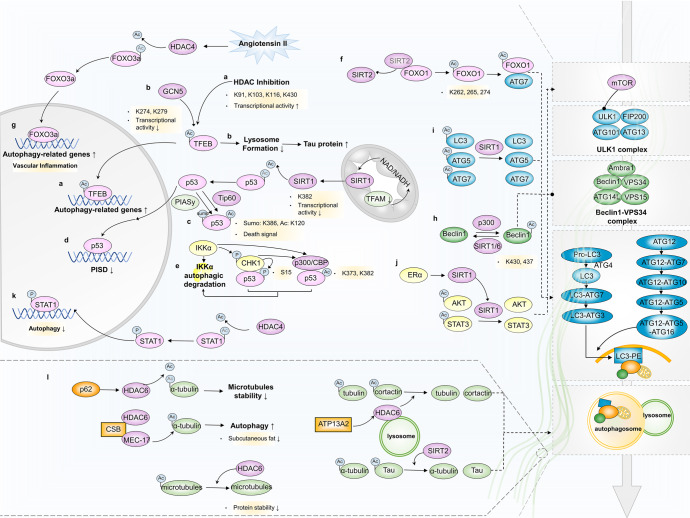
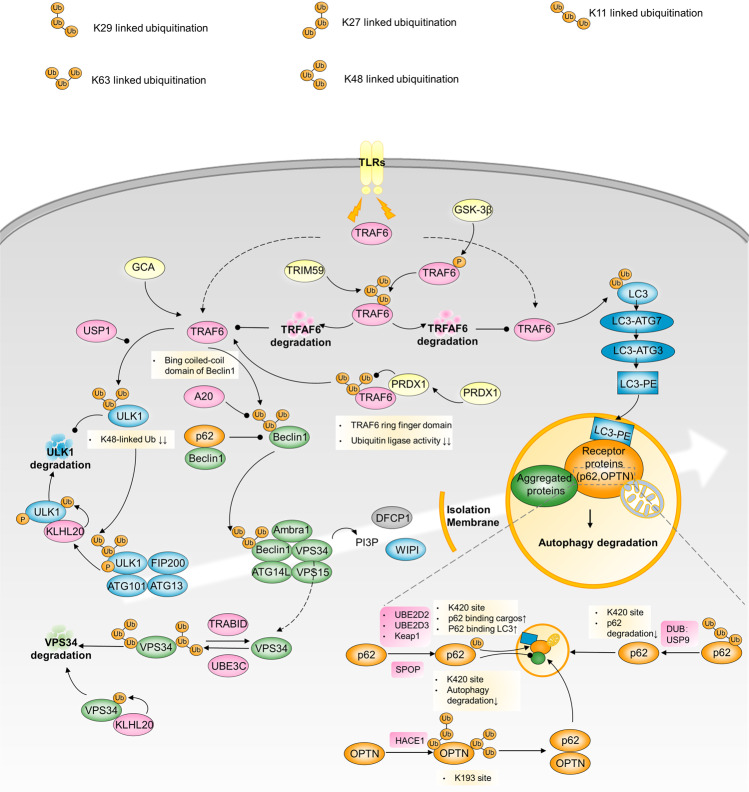
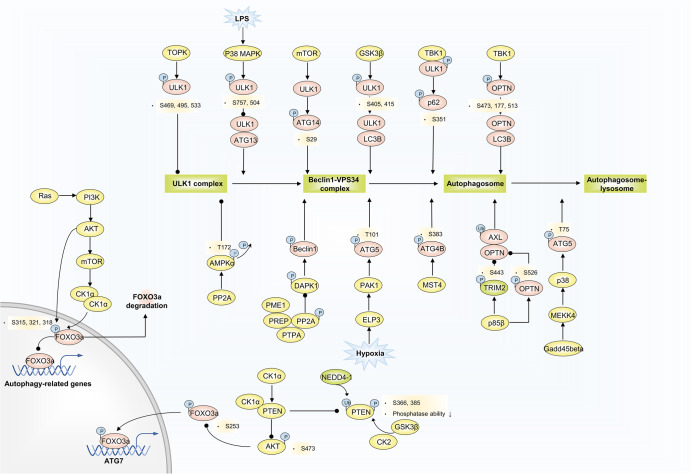
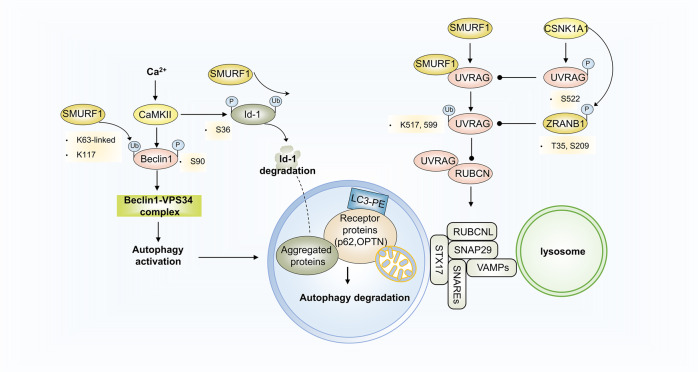
Autophagy is a conserved lysosomal degradation pathway where cellular components are dynamically degraded and re-processed to maintain physical homeostasis. However, the physiological effect of autophagy appears to be multifaced. On the one hand, autophagy functions as a cytoprotective mechanism, protecting against multiple diseases, especially tumor, cardiovascular disorders, and neurodegenerative and infectious disease. Conversely, autophagy may also play a detrimental role via pro-survival effects on cancer cells or cell-killing effects on normal body cells. During disorder onset and progression, the expression levels of autophagy-related regulators and proteins encoded by autophagy-related genes (ATGs) are abnormally regulated, giving rise to imbalanced autophagy flux. However, the detailed mechanisms and molecular events of this process are quite complex. Epigenetic, including DNA methylation, histone modifications and miRNAs, and post-translational modifications, including ubiquitination, phosphorylation and acetylation, precisely manipulate gene expression and protein function, and are strongly correlated with the occurrence and development of multiple diseases. There is substantial evidence that autophagy-relevant regulators and machineries are subjected to epigenetic and post-translational modulation, resulting in alterations in autophagy levels, which subsequently induces disease or affects the therapeutic effectiveness to agents. In this review, we focus on the regulatory mechanisms mediated by epigenetic and post-translational modifications in disease-related autophagy to unveil potential therapeutic targets. In addition, the effect of autophagy on the therapeutic effectiveness of epigenetic drugs or drugs targeting post-translational modification have also been discussed, providing insights into the combination with autophagy activators or inhibitors in the treatment of clinical diseases.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
