

Lysine-specific histone demethylase 1A (KDM1A/LSD1) inhibition attenuates DNA double-strand break repair and augments the efficacy of temozolomide in glioblastoma
- PMID: 36652263
- PMCID: PMC10326496
- DOI: 10.1093/neuonc/noad018
Lysine-specific histone demethylase 1A (KDM1A/LSD1) inhibition attenuates DNA double-strand break repair and augments the efficacy of temozolomide in glioblastoma
Abstract
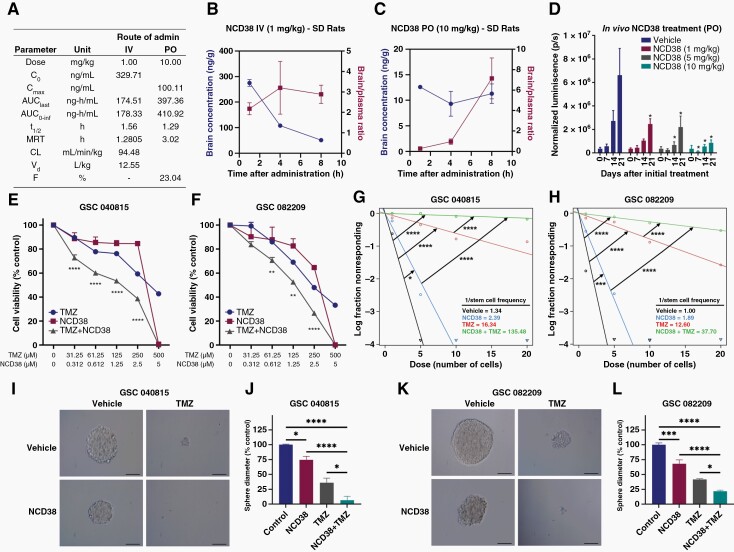
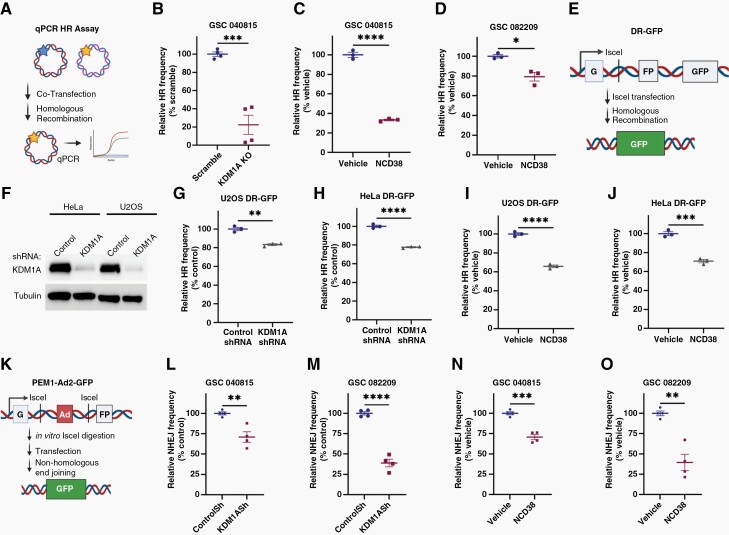
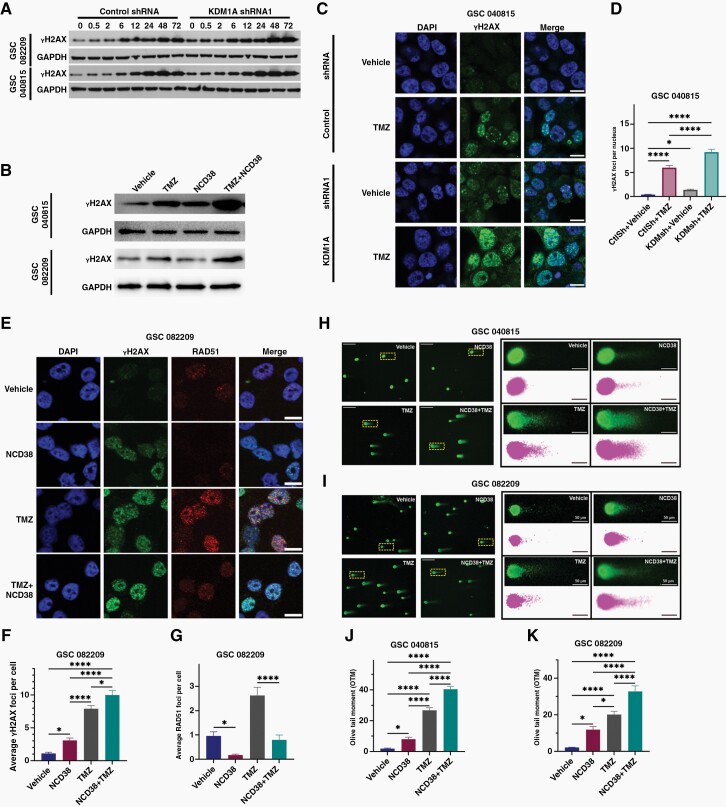
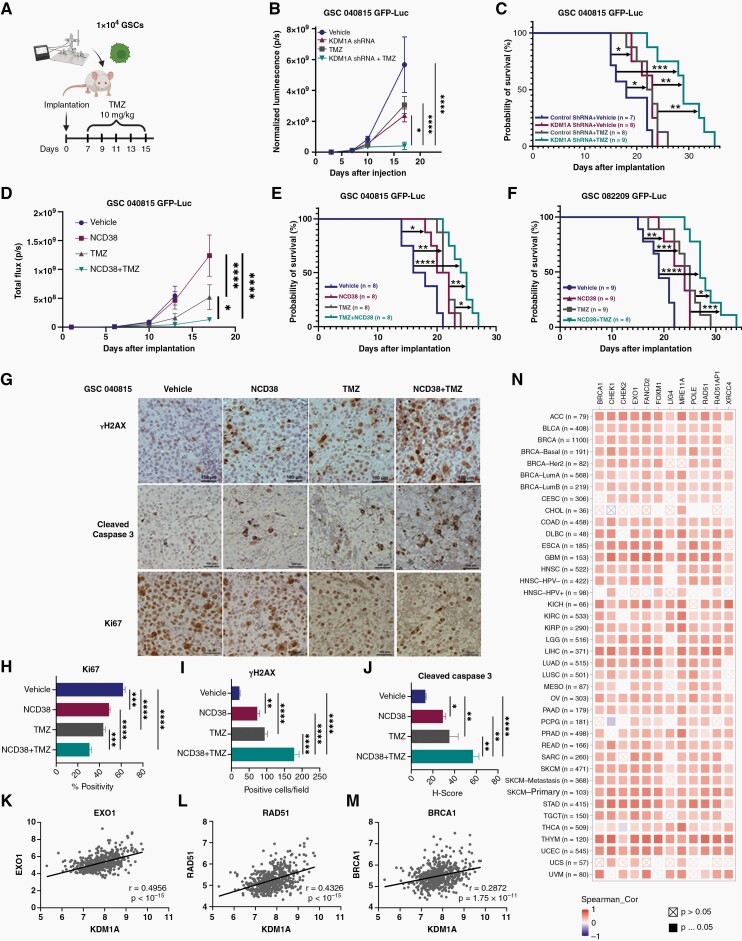
Background: Efficient DNA repair in response to standard chemo and radiation therapies often contributes to glioblastoma (GBM) therapy resistance. Understanding the mechanisms of therapy resistance and identifying the drugs that enhance the therapeutic efficacy of standard therapies may extend the survival of GBM patients. In this study, we investigated the role of KDM1A/LSD1 in DNA double-strand break (DSB) repair and a combination of KDM1A inhibitor and temozolomide (TMZ) in vitro and in vivo using patient-derived glioma stem cells (GSCs).
Methods: Brain bioavailability of the KDM1A inhibitor (NCD38) was established using LS-MS/MS. The effect of a combination of KDM1A knockdown or inhibition with TMZ was studied using cell viability and self-renewal assays. Mechanistic studies were conducted using CUT&Tag-seq, RNA-seq, RT-qPCR, western blot, homologous recombination (HR) and non-homologous end joining (NHEJ) reporter, immunofluorescence, and comet assays. Orthotopic murine models were used to study efficacy in vivo.
Results: TCGA analysis showed KDM1A is highly expressed in TMZ-treated GBM patients. Knockdown or knockout or inhibition of KDM1A enhanced TMZ efficacy in reducing the viability and self-renewal of GSCs. Pharmacokinetic studies established that NCD38 readily crosses the blood-brain barrier. CUT&Tag-seq studies showed that KDM1A is enriched at the promoters of DNA repair genes and RNA-seq studies confirmed that KDM1A inhibition reduced their expression. Knockdown or inhibition of KDM1A attenuated HR and NHEJ-mediated DNA repair capacity and enhanced TMZ-mediated DNA damage. A combination of KDM1A knockdown or inhibition and TMZ treatment significantly enhanced the survival of tumor-bearing mice.
Conclusions: Our results provide evidence that KDM1A inhibition sensitizes GBM to TMZ via attenuation of DNA DSB repair pathways.
Keywords: DNA repair; KDM1A/LSD1; glioblastoma; glioma stem cells; temozolomide.
© The Author(s) 2023. Published by Oxford University Press on behalf of the Society for Neuro-Oncology. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Conflict of interest statement
None.
Figures


References
-
- Omuro A, DeAngelis LM. Glioblastoma and other malignant gliomas: a clinical review. JAMA. 2013;310(17):1842–1850. - PubMed
-
- Johnson DR, O’Neill BP. Glioblastoma survival in the United States before and during the temozolomide era. J Neurooncol. 2012;107(2):359–364. - PubMed
-
- Cloughesy TF, Cavenee WK, Mischel PS. Glioblastoma: from molecular pathology to targeted treatment. Annu Rev Pathol. 2014;9:1–25. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
