

A molecular network of conserved factors keeps ribosomes dormant in the egg
- PMID: 36653451
- PMCID: PMC7614339
- DOI: 10.1038/s41586-022-05623-y
A molecular network of conserved factors keeps ribosomes dormant in the egg
Abstract
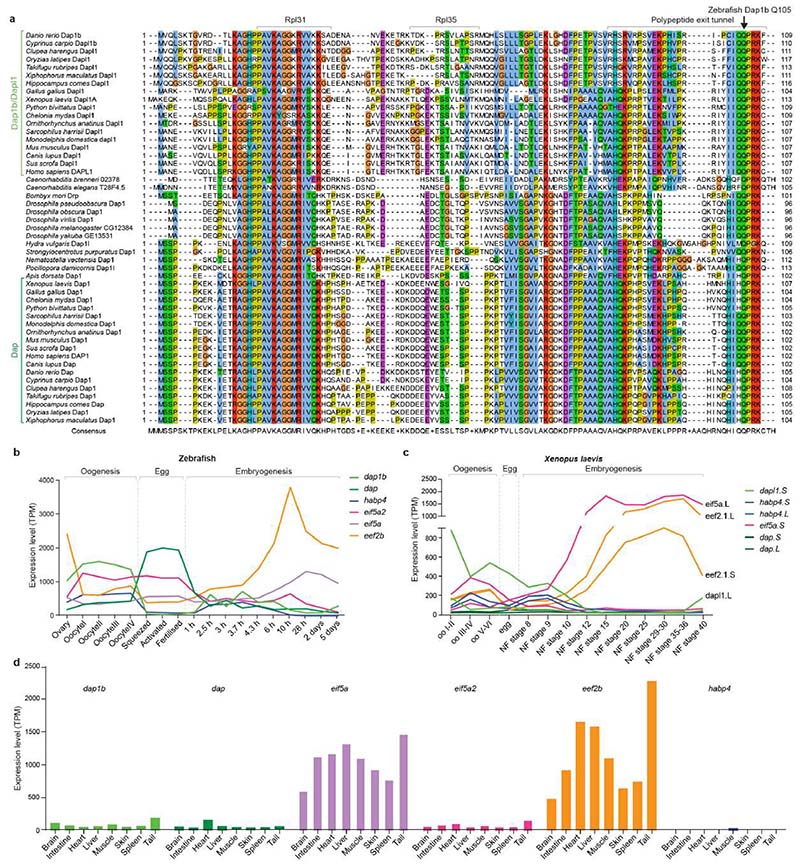
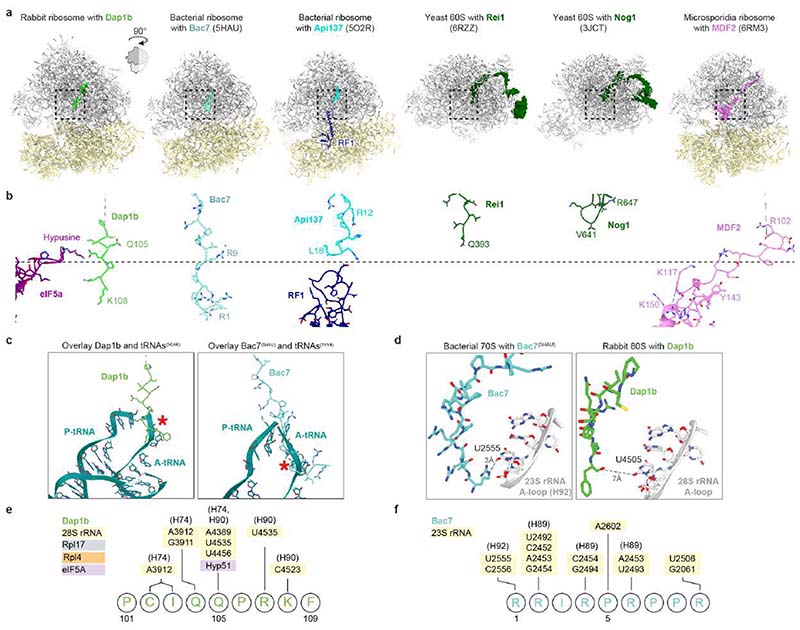
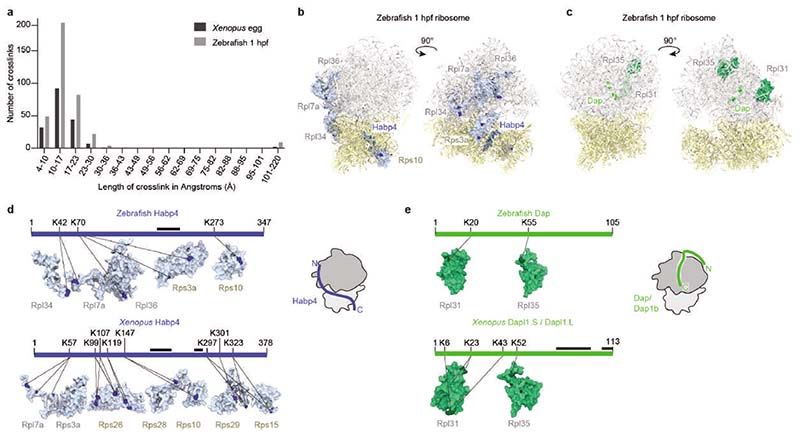
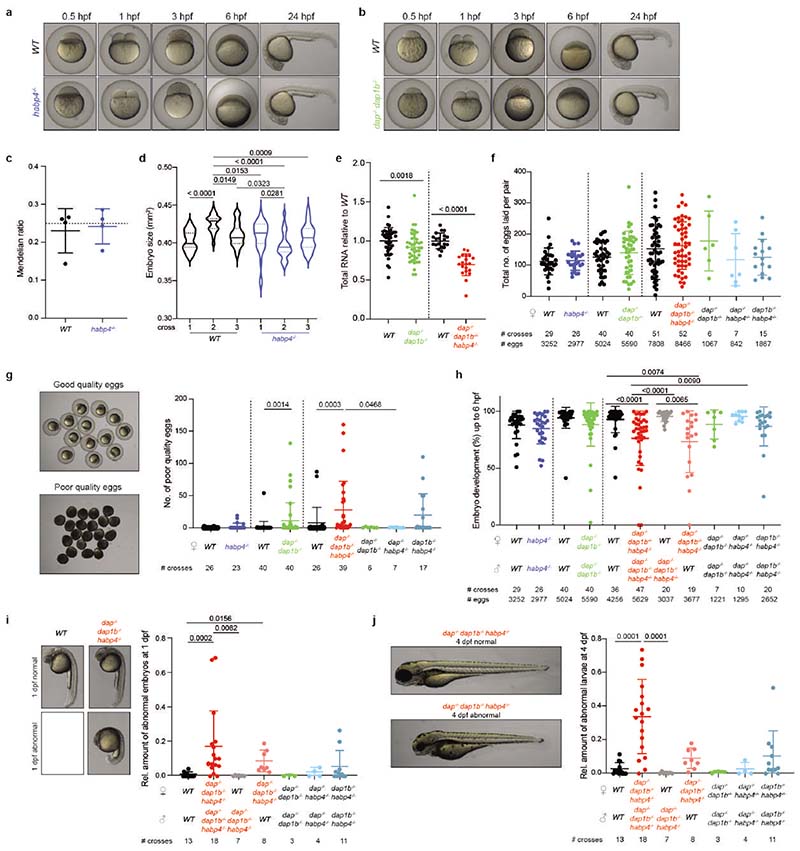
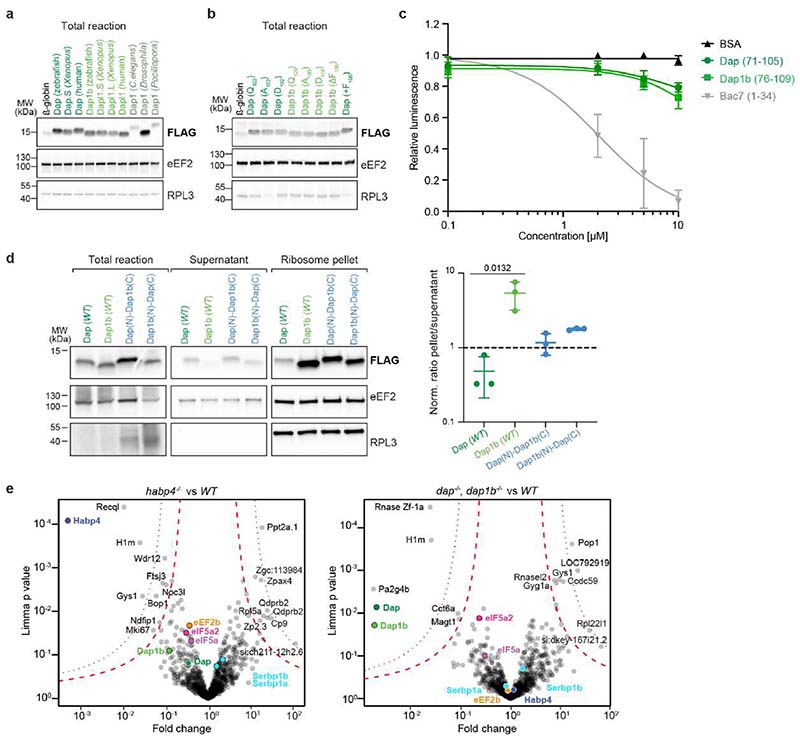
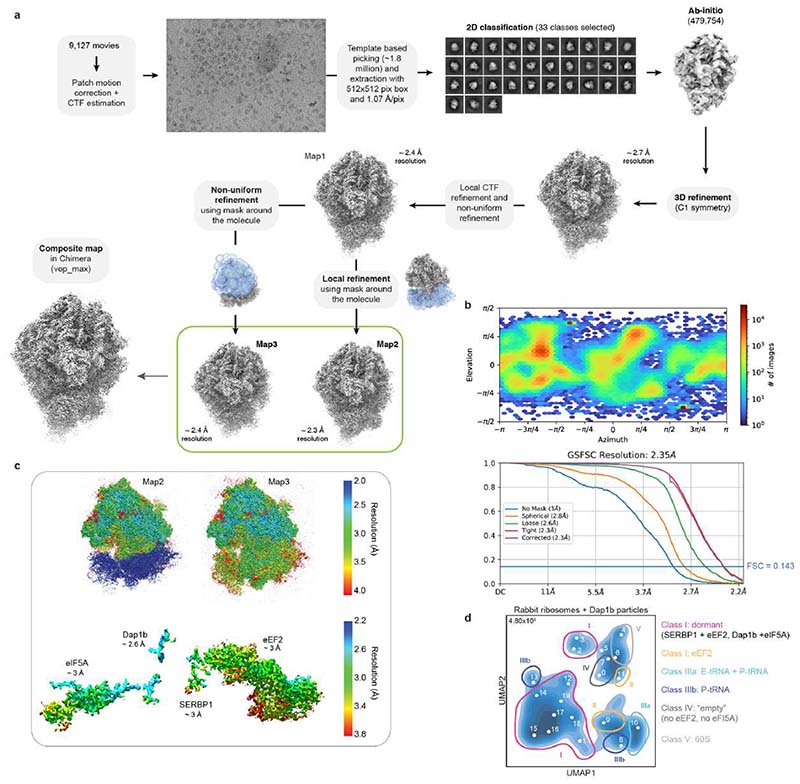
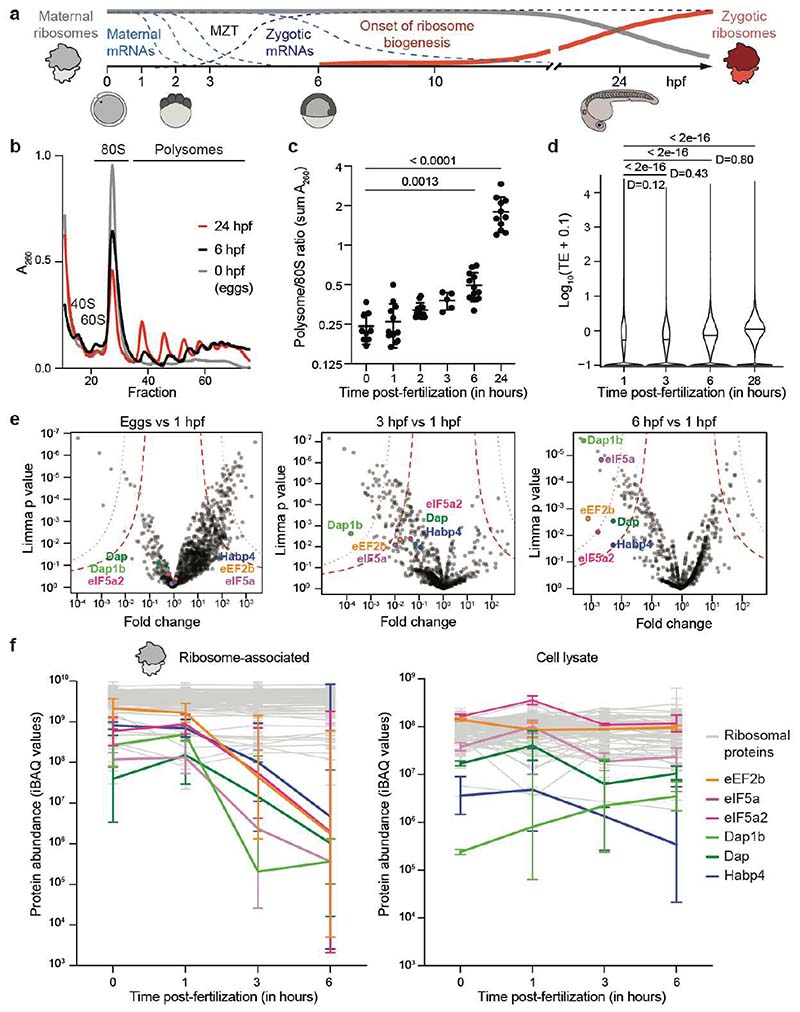
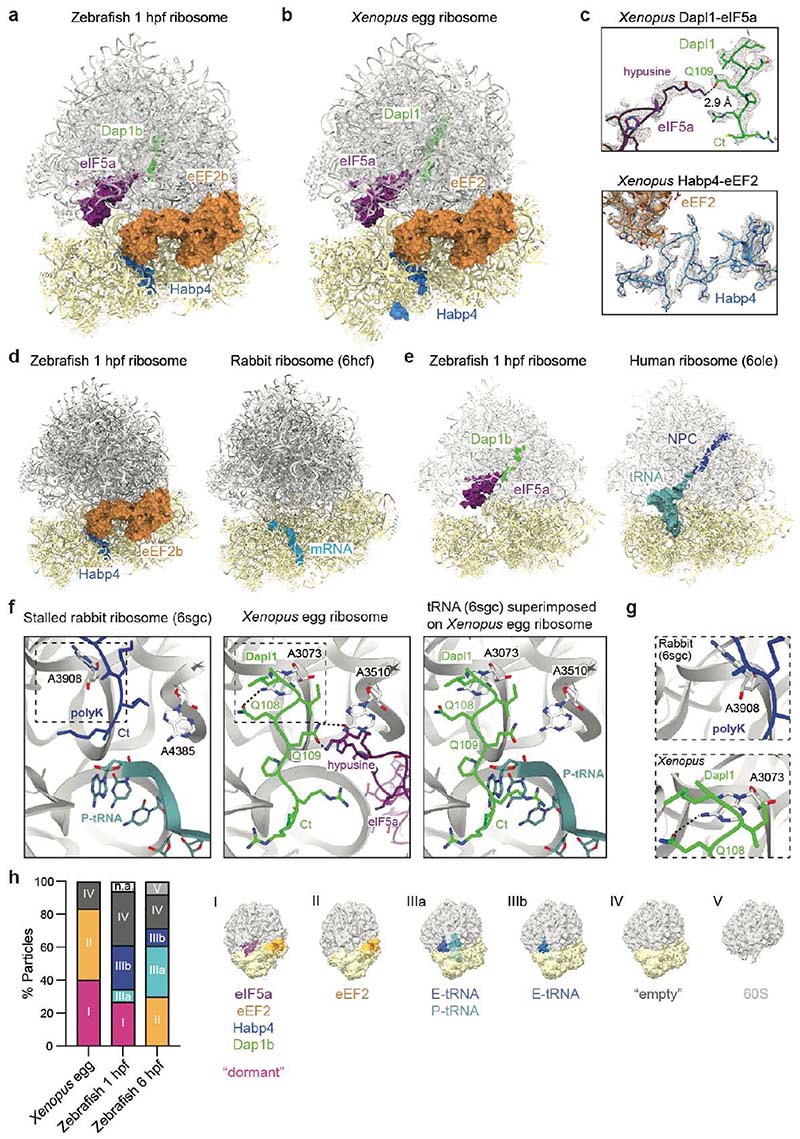
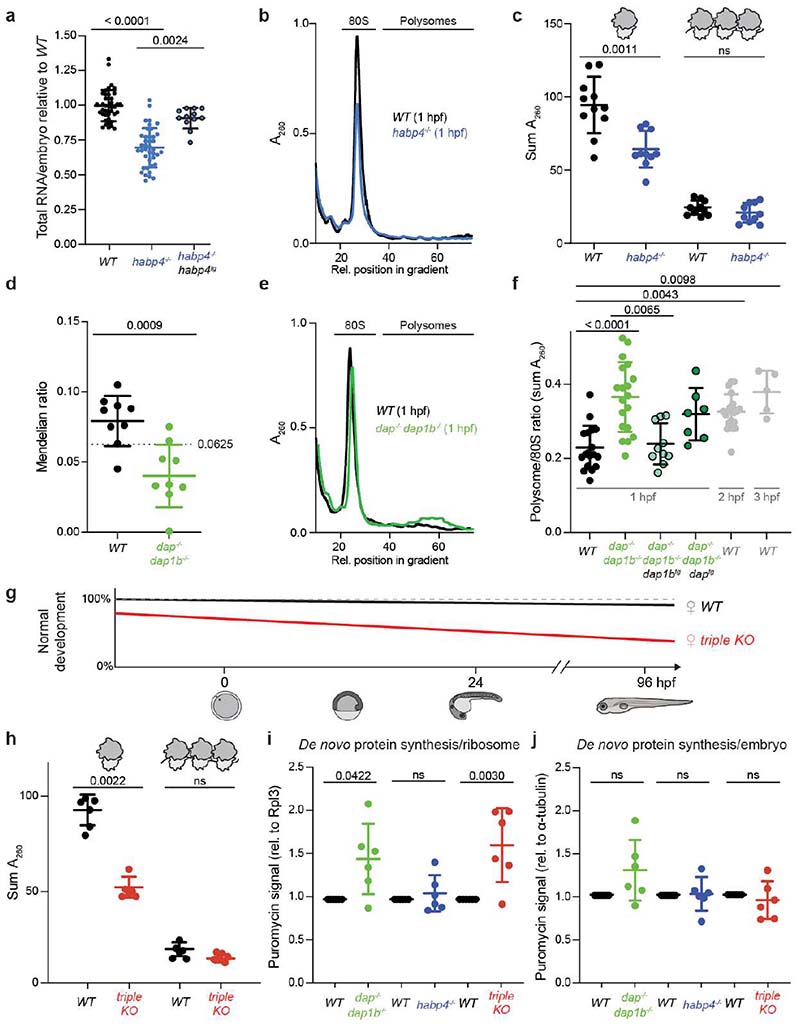
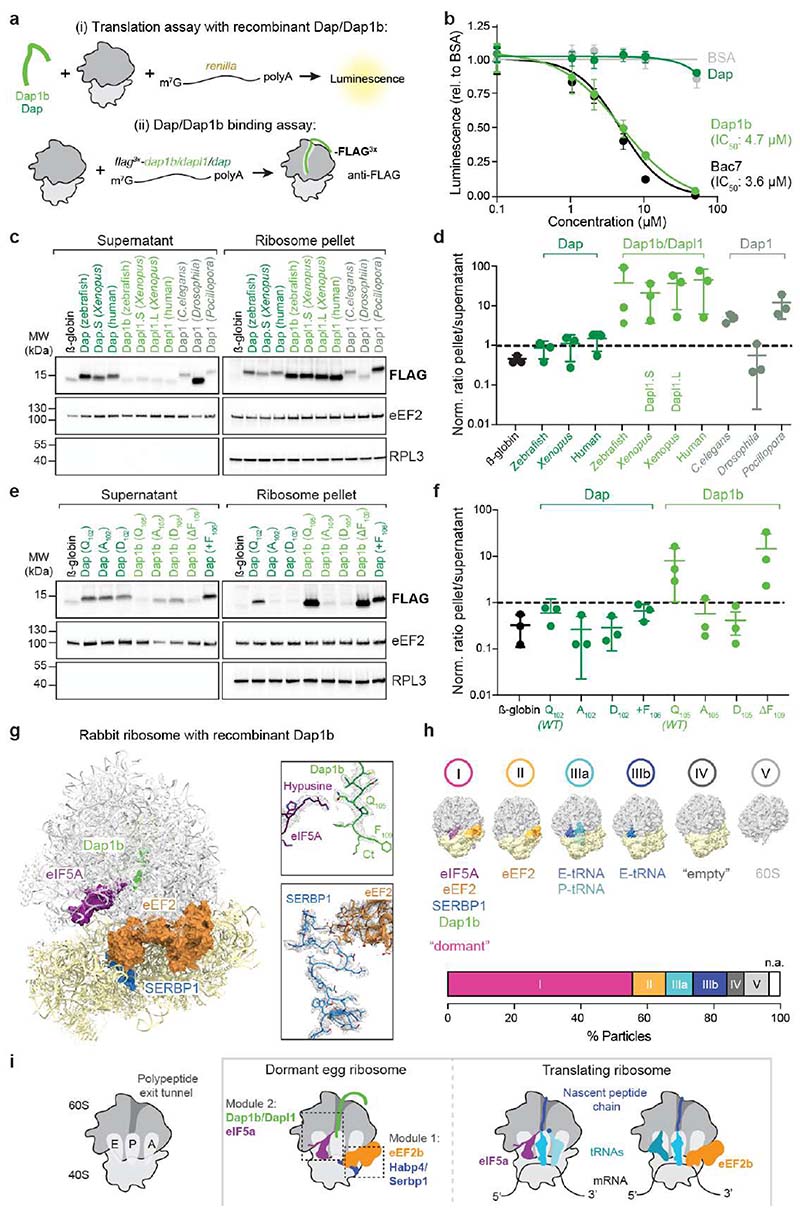
Ribosomes are produced in large quantities during oogenesis and are stored in the egg. However, the egg and early embryo are translationally repressed1-4. Here, using mass spectrometry and cryo-electron microscopy analyses of ribosomes isolated from zebrafish (Danio rerio) and Xenopus laevis eggs and embryos, we provide molecular evidence that ribosomes transition from a dormant state to an active state during the first hours of embryogenesis. Dormant ribosomes are associated with four conserved factors that form two modules, consisting of Habp4-eEF2 and death associated protein 1b (Dap1b) or Dap in complex with eIF5a. Both modules occupy functionally important sites and act together to stabilize ribosomes and repress translation. Dap1b (also known as Dapl1 in mammals) is a newly discovered translational inhibitor that stably inserts into the polypeptide exit tunnel. Addition of recombinant zebrafish Dap1b protein is sufficient to block translation and reconstitute the dormant egg ribosome state in a mammalian translation extract in vitro. Thus, a developmentally programmed, conserved ribosome state has a key role in ribosome storage and translational repression in the egg.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Figures
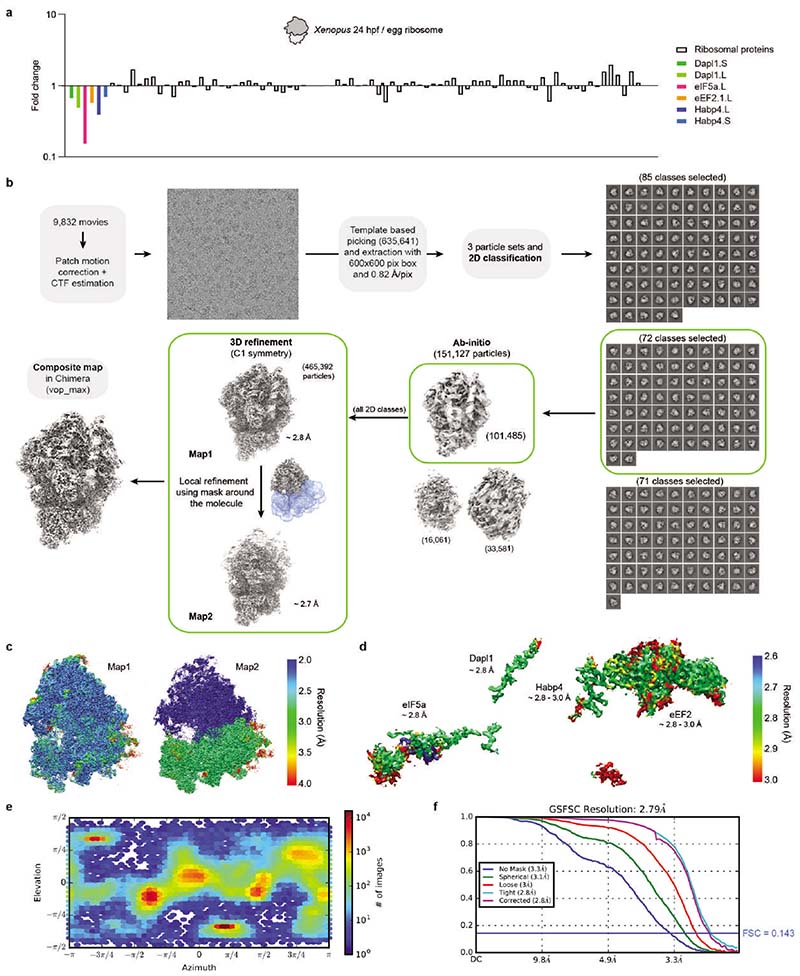
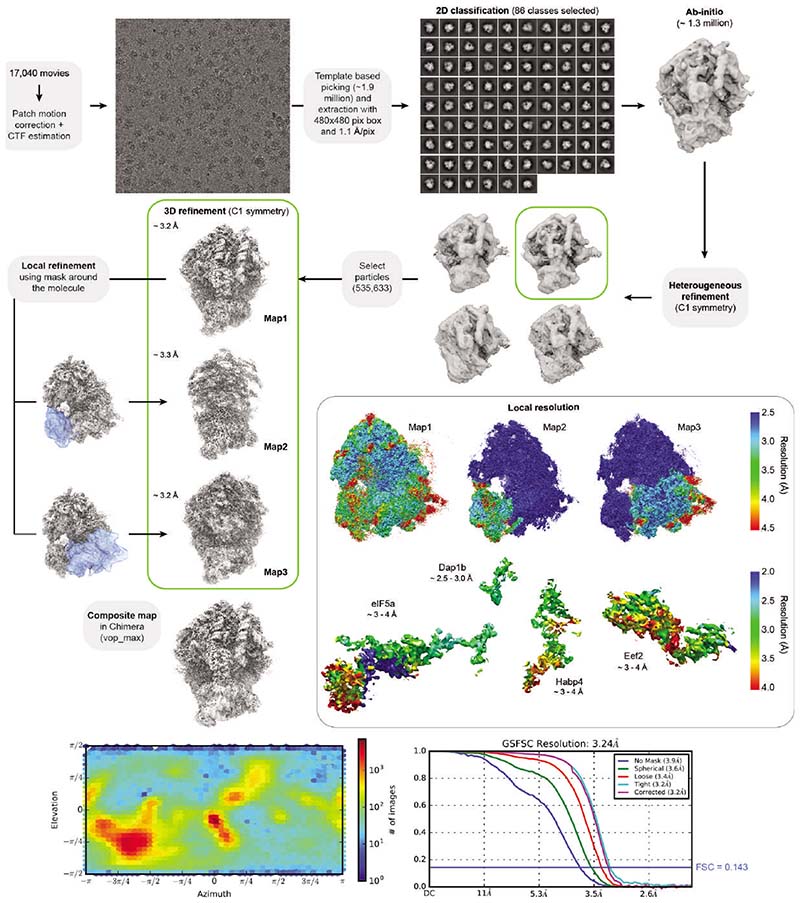
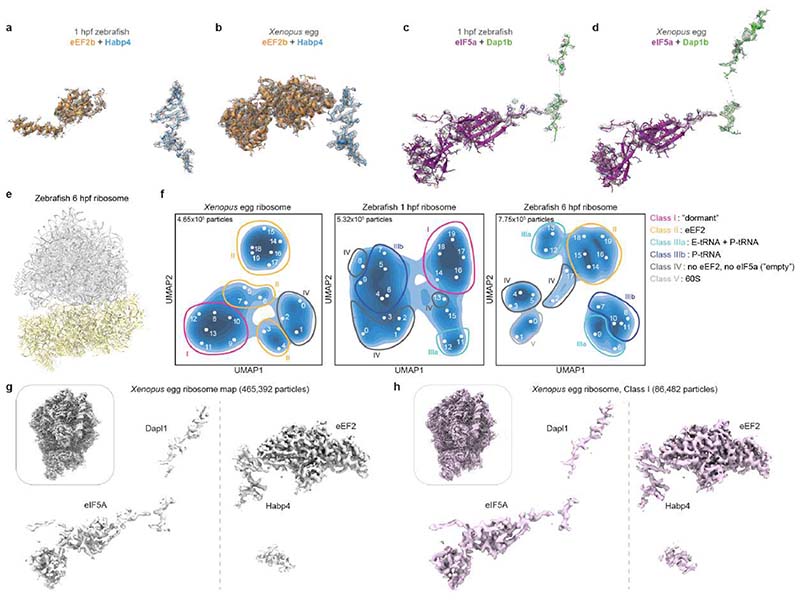
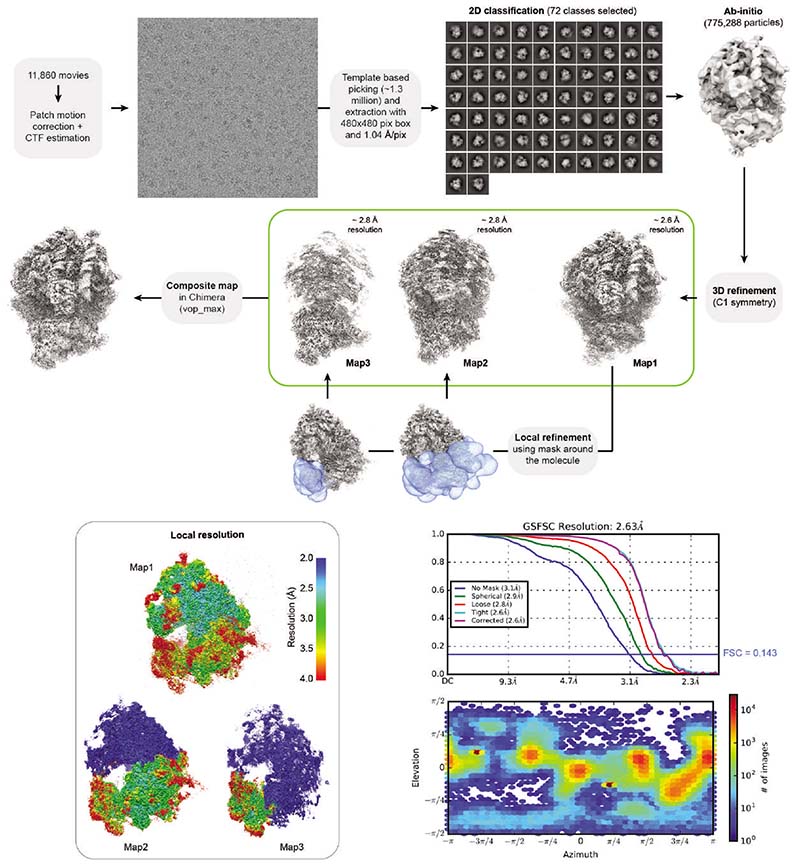
References
-
- Woodland HR. Changes in the polysome content of developing Xenopus laevis embryos. Developmental Biology. 1974;40:90–101. - PubMed
-
- Brandis JW, Raff RA. Translation of oogenetic mRNA in sea urchin eggs and early embryos. Demonstration of a change in translational efficiency following fertilization. Dev Biol. 1978;67:99–113. - PubMed
-
- Bachvarova R, De Leon V. Stored and polysomal ribosomes of mouse ova. Developmental Biology. 1977;58:248–254. - PubMed
-
- Burkholder GD, Comings DE, Okada TA. A storage form of ribosomes in mouse oocytes. Experimental Cell Research. 1971;69:361–371. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
