

Chromodomain on Y-like 2 (CDYL2) implicated in mitosis and genome stability regulation via interaction with CHAMP1 and POGZ
- PMID: 36658409
- PMCID: PMC11072993
- DOI: 10.1007/s00018-022-04659-7
Chromodomain on Y-like 2 (CDYL2) implicated in mitosis and genome stability regulation via interaction with CHAMP1 and POGZ
Abstract
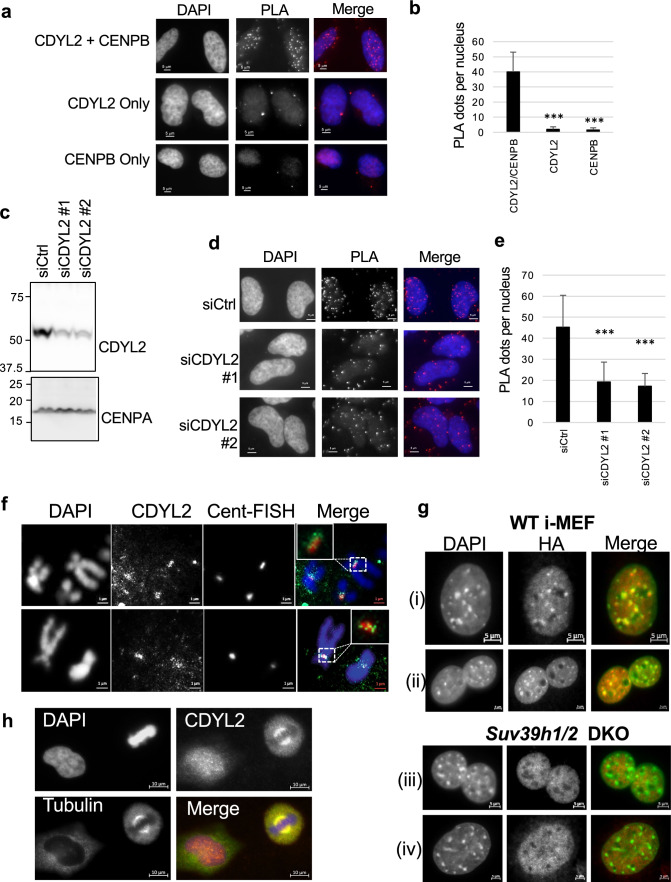
Histone H3 trimethylation on lysine 9 (H3K9me3) is a defining feature of mammalian pericentromeres, loss of which results in genome instability. Here we show that CDYL2 is recruited to pericentromeres in an H3K9me3-dependent manner and is required for faithful mitosis and genome stability. CDYL2 RNAi in MCF-7 breast cancer cells and Hela cervical cancer cells inhibited their growth, induced apoptosis, and provoked both nuclear and mitotic aberrations. Mass spectrometry analysis of CDYL2-interacting proteins identified the neurodevelopmental disease-linked mitotic regulators CHAMP1 and POGZ, which are associated with a central non-conserved region of CDYL2. RNAi rescue assays identified both the CDYL2 chromodomain and the CHAMP1-POGZ interacting region as required and, together, sufficient for CDYL2 regulation of mitosis and genome stability. CDYL2 RNAi caused loss of CHAMP1 localization at pericentromeres. We propose that CDYL2 functions as an adaptor protein that connects pericentromeric H3K9me3 with CHAMP1 and POGZ to ensure mitotic fidelity and genome stability.
Keywords: C13orf8; CAMP; Centromere; Epigenetics; ZNF828.
© 2023. The Author(s), under exclusive licence to Springer Nature Switzerland AG.
Conflict of interest statement
The authors have no competing interests to declare.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
