

Closing the gap to effective gene drive in Aedes aegypti by exploiting germline regulatory elements
- PMID: 36670107
- PMCID: PMC9860013
- DOI: 10.1038/s41467-023-36029-7
Closing the gap to effective gene drive in Aedes aegypti by exploiting germline regulatory elements
Abstract
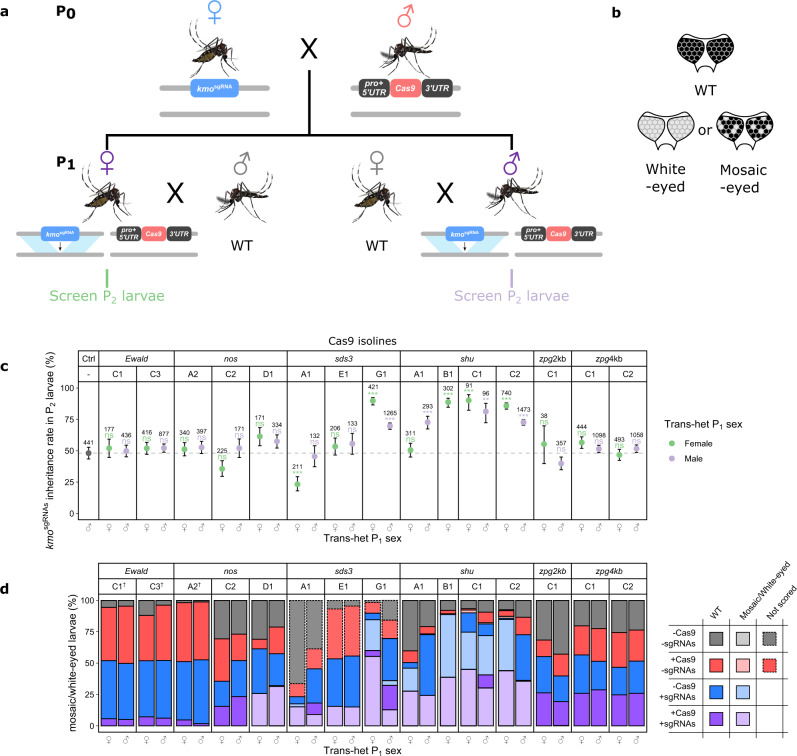
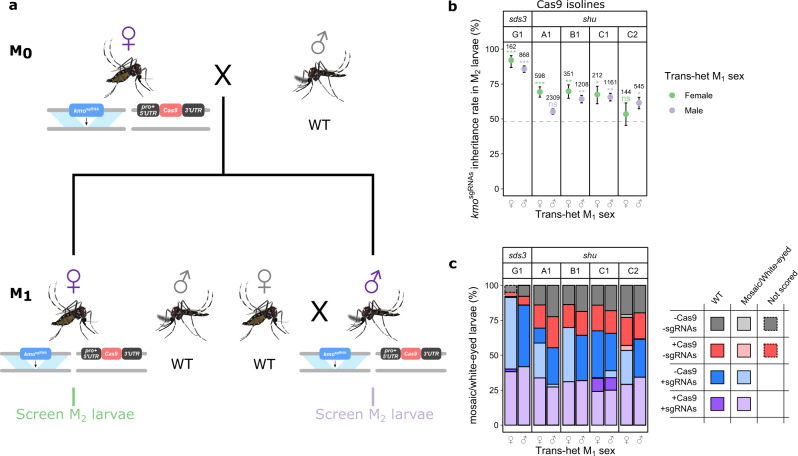
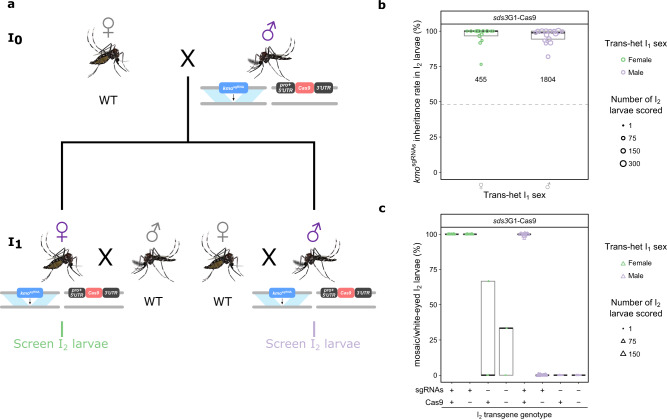
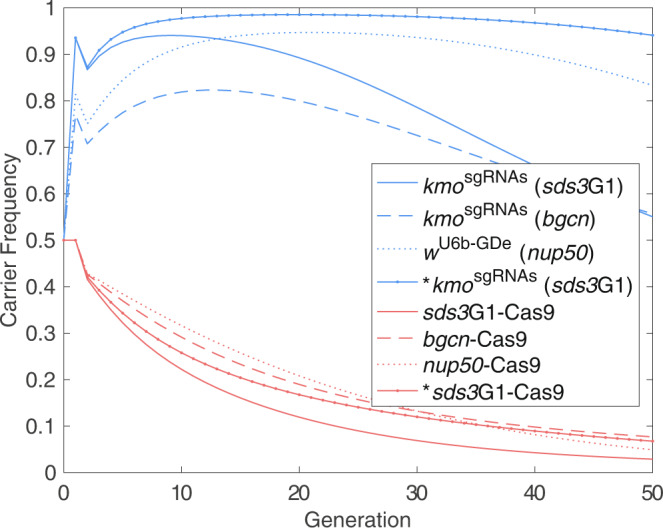
CRISPR/Cas9-based homing gene drives have emerged as a potential new approach to mosquito control. While attempts have been made to develop such systems in Aedes aegypti, none have been able to match the high drive efficiency observed in Anopheles species. Here we generate Ae. aegypti transgenic lines expressing Cas9 using germline-specific regulatory elements and assess their ability to bias inheritance of an sgRNA-expressing element (kmosgRNAs). Four shu-Cas9 and one sds3-Cas9 isolines can significantly bias the inheritance of kmosgRNAs, with sds3G1-Cas9 causing the highest average inheritance of ~86% and ~94% from males and females carrying both elements outcrossed to wild-type, respectively. Our mathematical model demonstrates that sds3G1-Cas9 could enable the spread of the kmosgRNAs element to either reach a higher (by ~15 percentage point) maximum carrier frequency or to achieve similar maximum carrier frequency faster (by 12 generations) when compared to two other established split drive systems.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
- BB/M011224/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/I/00007038/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 110117/Z/15/Z/WT_/Wellcome Trust/United Kingdom
- BBS/E/I/00007039/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Miscellaneous
