

Novel Ground-Up 3D Multicellular Simulators for Synthetic Biology CAD Integrating Stochastic Gillespie Simulations Benchmarked with Topologically Variable SBML Models
- PMID: 36672895
- PMCID: PMC9859520
- DOI: 10.3390/genes14010154
Novel Ground-Up 3D Multicellular Simulators for Synthetic Biology CAD Integrating Stochastic Gillespie Simulations Benchmarked with Topologically Variable SBML Models
Abstract
The elevation of Synthetic Biology from single cells to multicellular simulations would be a significant scale-up. The spatiotemporal behavior of cellular populations has the potential to be prototyped in silico for computer assisted design through ergonomic interfaces. Such a platform would have great practical potential across medicine, industry, research, education and accessible archiving in bioinformatics. Existing Synthetic Biology CAD systems are considered limited regarding population level behavior, and this work explored the in silico challenges posed from biological and computational perspectives. Retaining the connection to Synthetic Biology CAD, an extension of the Infobiotics Workbench Suite was considered, with potential for the integration of genetic regulatory models and/or chemical reaction networks through Next Generation Stochastic Simulator (NGSS) Gillespie algorithms. These were executed using SBML models generated by in-house SBML-Constructor over numerous topologies and benchmarked in association with multicellular simulation layers. Regarding multicellularity, two ground-up multicellular solutions were developed, including the use of Unreal Engine 4 contrasted with CPU multithreading and Blender visualization, resulting in a comparison of real-time versus batch-processed simulations. In conclusion, high-performance computing and client-server architectures could be considered for future works, along with the inclusion of numerous biologically and physically informed features, whilst still pursuing ergonomic solutions.
Keywords: CAD; SBML; automation; biophysics; chemical reaction networks; multicellular simulation; stochastic Gillespie; synthetic biology; systems biology; unreal engine.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses or interpretation of the data; in the writing of the manuscript or in the decision to publish the results.
Figures







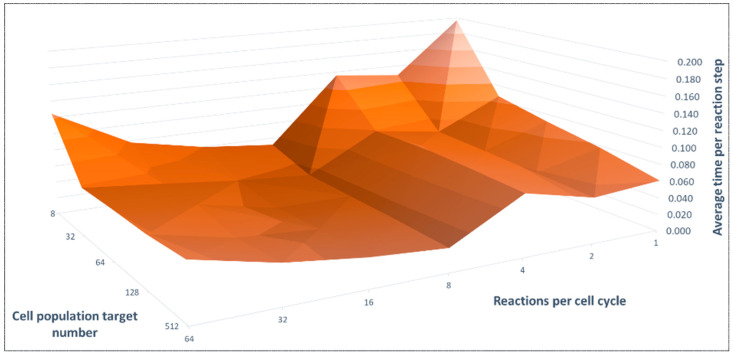



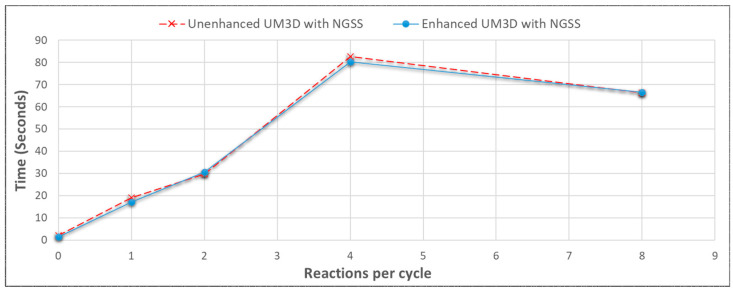




References
-
- Mirams G.R., Arthurs C.J., Bernabeu M.O., Bordas R., Cooper J., Corrias A., Davit Y., Dunn S.J., Fletcher A.G., Harvey D.G., et al. Chaste: An open source C++ library for computational physiology and biology. PLoS Comput. Biol. 2013;9:e1002970. doi: 10.1371/journal.pcbi.1002970. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
