

Estradiol and Estrone Have Different Biological Functions to Induce NF-κB-Driven Inflammation, EMT and Stemness in ER+ Cancer Cells
- PMID: 36674737
- PMCID: PMC9865376
- DOI: 10.3390/ijms24021221
Estradiol and Estrone Have Different Biological Functions to Induce NF-κB-Driven Inflammation, EMT and Stemness in ER+ Cancer Cells
Abstract
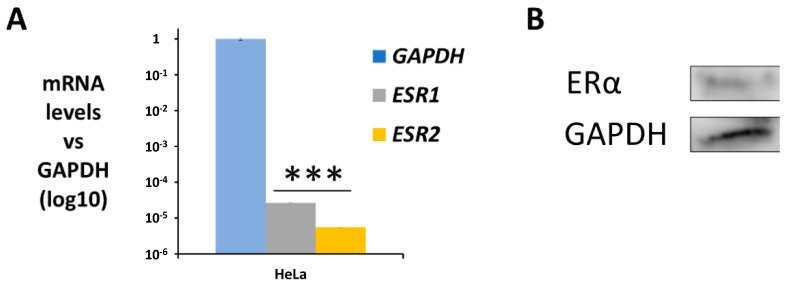
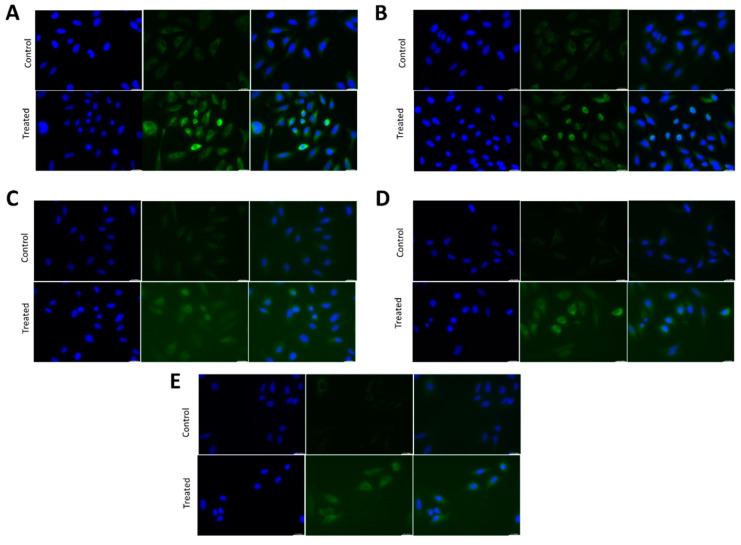
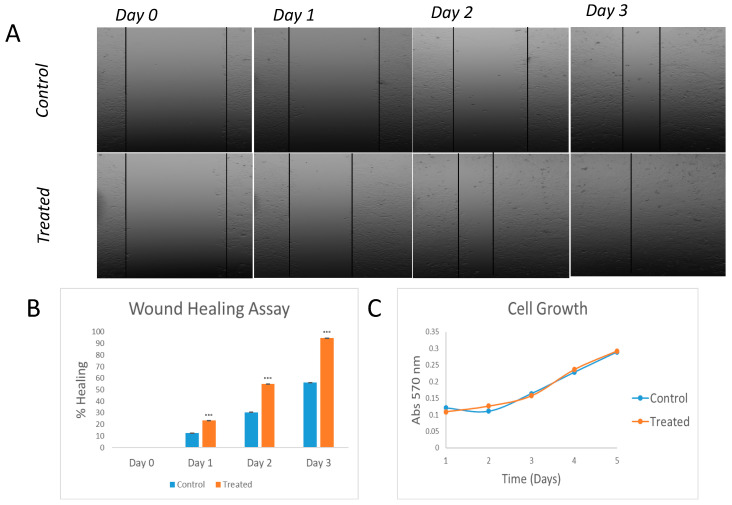
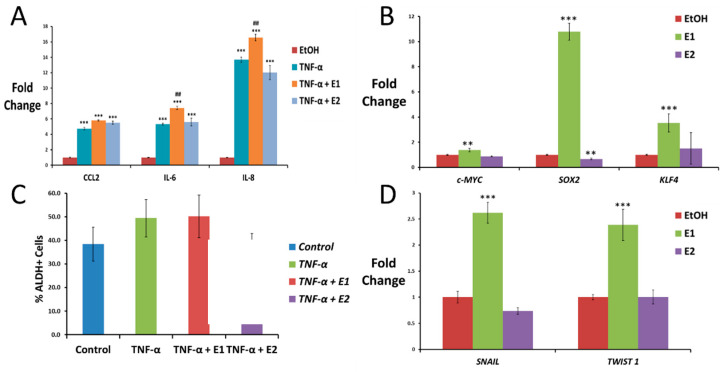
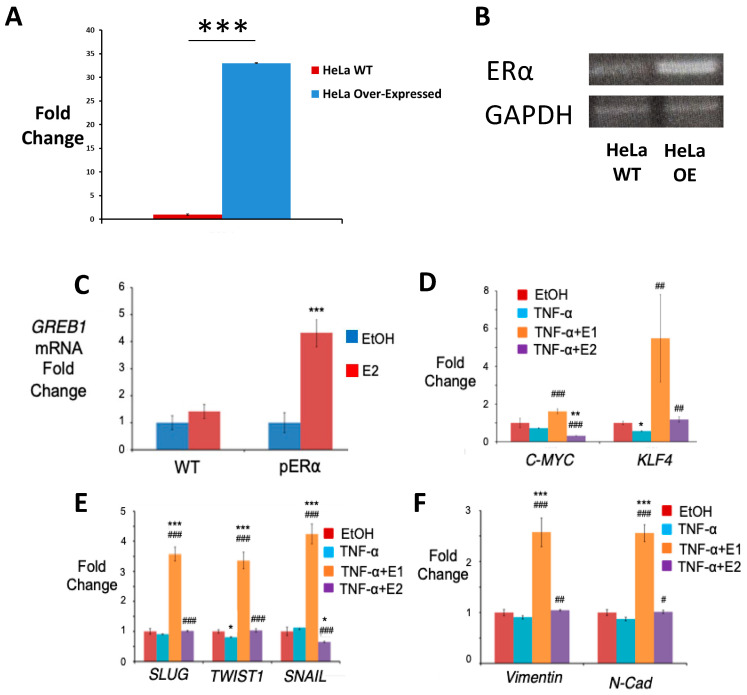
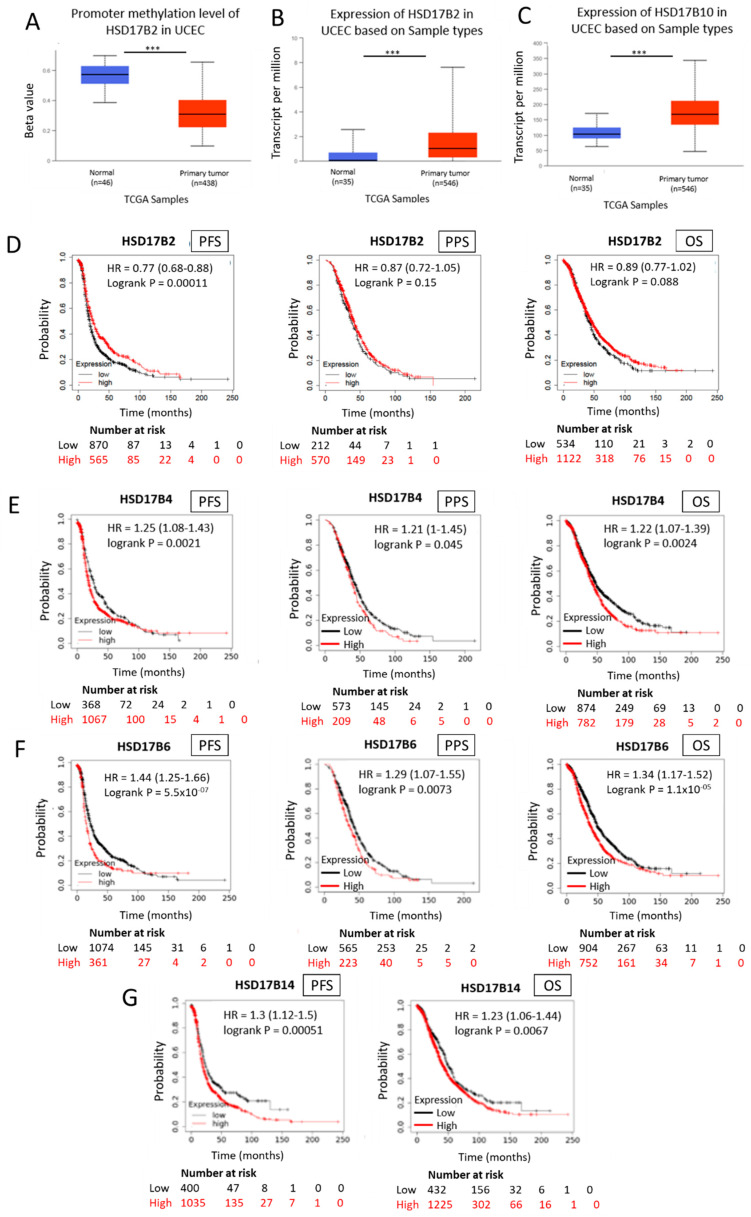
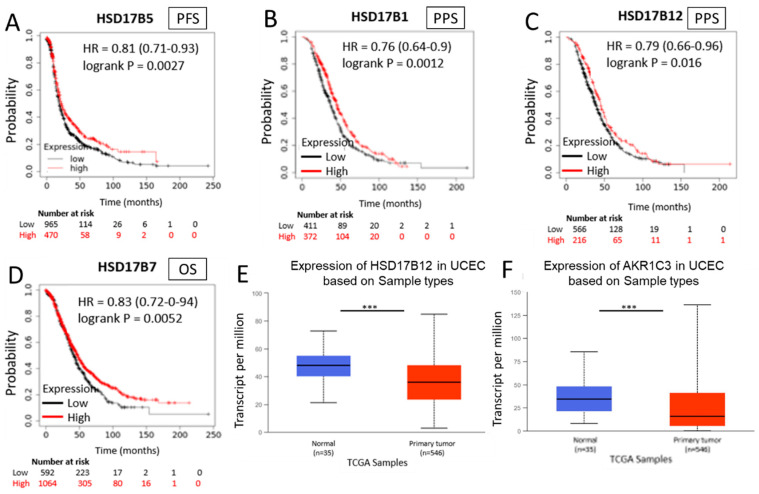
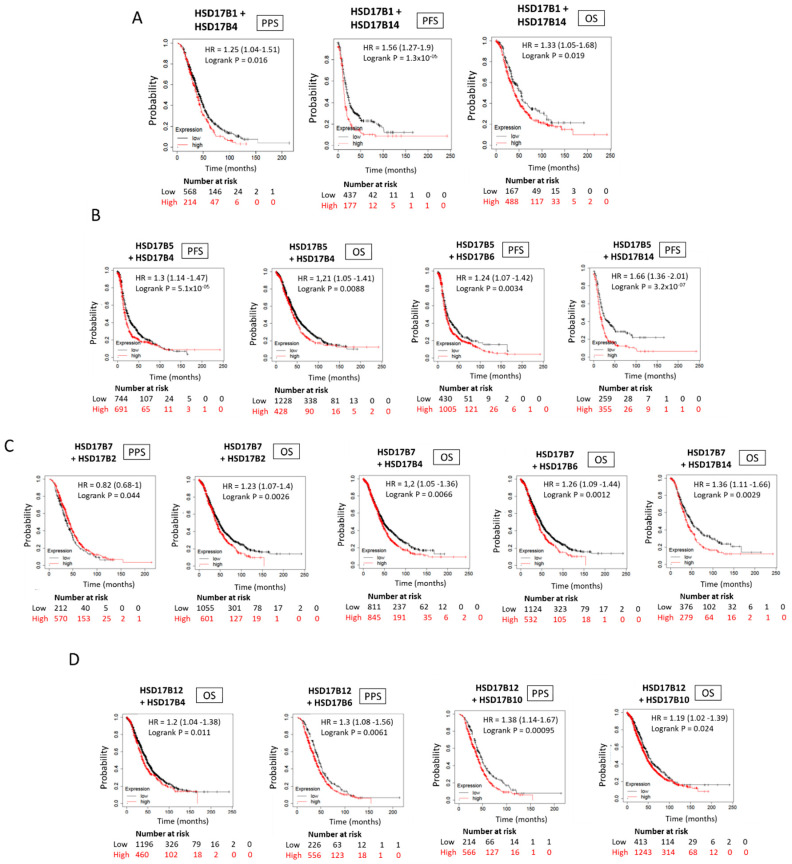
In general, the risk of being diagnosed with cancer increases with age; however, the development of estrogen-receptor-positive (ER+) cancer types in women are more closely related to menopausal status than age. In fact, the general risk factors for cancer development, such as obesity-induced inflammation, show differences in their association with ER+ cancer risk in pre- and postmenopausal women. Here, we tested the role of the principal estrogens in the bloodstream before and after menopause, estradiol (E2) and estrone (E1), respectively, on inflammation, epithelial-to-mesenchymal transition (EMT) and cancer stem cell enrichment in the human ER+ cervical cancer cell line HeLa. Our results demonstrate that E1, contrary to E2, is pro-inflammatory, increases embryonic stem-transcription factors (ES-TFs) expression and induces EMT in ER+ HeLa cells. Moreover, we observed that high intratumoural expression levels of 17β-Hydroxysteroid dehydrogenase (HSD17B) isoforms involved in E1 synthesis is a poor prognosis factor, while overexpression of E2-synthetizing HSD17B isoforms is associated with a better outcome, for patients diagnosed with ER+ ovarian and uterine corpus carcinomas. This work demonstrates that E1 and E2 have different biological functions in ER+ gynaecologic cancers. These results open a new line of research in the study of ER+ cancer subtypes, highlighting the potential key oncogenic role of E1 and HSD17B E1-synthesizing enzymes in the development and progression of these diseases.
Keywords: ER+ cancer; HSD17B; NF-κB; estradiol; estrone; inflammation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Substances
Grants and funding
- PIE16-00045/Instituto de Salud Carlos III
- PEMP-0205-2020/Ministry of Health and Family of the Junta de Andalucía and European Regional Development Fund
- AEI/10.13039/501100011033/FEDER/Spanish Ministry of Science, Innovation and Universities
- RTI2018-101309-B-C22/"Una manera de hacer Europa"
- MSCA-IF-2018 845104/European Commission
LinkOut - more resources
Full Text Sources
Medical
