

Effects of Sequence Composition, Patterning and Hydrodynamics on the Conformation and Dynamics of Intrinsically Disordered Proteins
- PMID: 36674958
- PMCID: PMC9867189
- DOI: 10.3390/ijms24021444
Effects of Sequence Composition, Patterning and Hydrodynamics on the Conformation and Dynamics of Intrinsically Disordered Proteins
Abstract
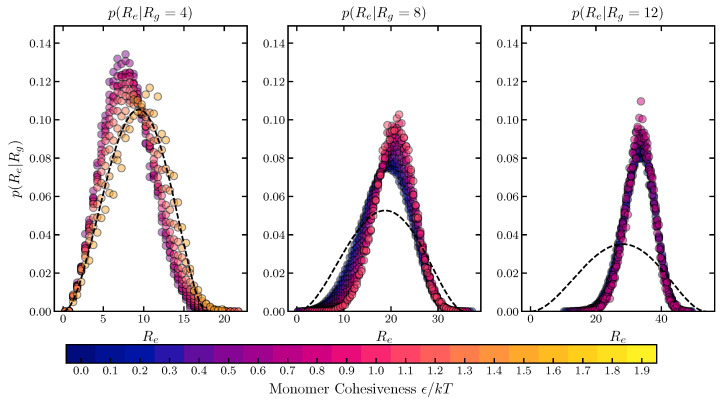
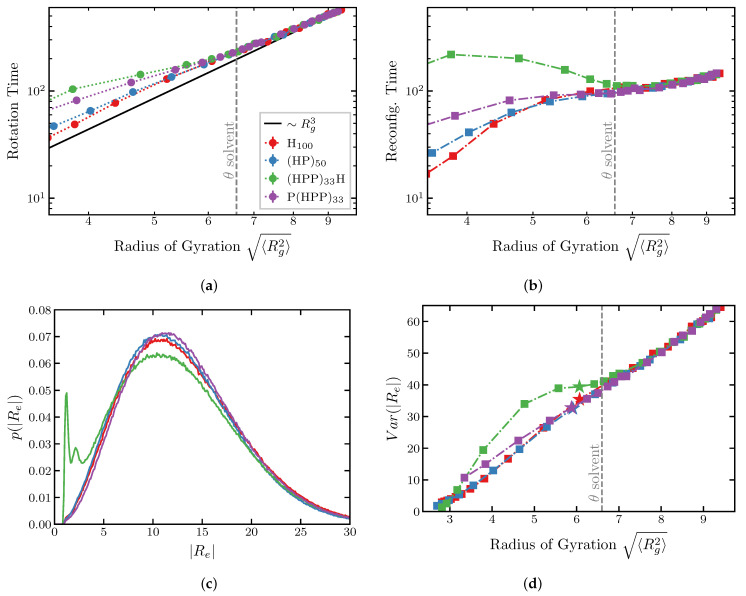
Intrinsically disordered proteins (IDPs) and intrinsically disordered regions (IDRs) perform diverse functions in cellular organization, transport and signaling. Unlike the well-defined structures of the classical natively folded proteins, IDPs and IDRs dynamically span large conformational and structural ensembles. This dynamic disorder impedes the study of the relationship between the amino acid sequences of the IDPs and their spatial structures and dynamics, with different experimental techniques often offering seemingly contradictory results. Although experimental and theoretical evidence indicates that some IDP properties can be understood based on their average biophysical properties and amino acid composition, other aspects of IDP function are dictated by the specifics of the amino acid sequence. We investigate the effects of several key variables on the dimensions and the dynamics of IDPs using coarse-grained polymer models. We focus on the sequence "patchiness" informed by the sequence and biophysical properties of different classes of IDPs-and in particular FG nucleoporins of the nuclear pore complex (NPC). We show that the sequence composition and patterning are well reflected in the global conformational variables such as the radius of gyration and hydrodynamic radius, while the end-to-end distance and dynamics are highly sequence-specific. We find that in good solvent conditions highly heterogeneous sequences of IDPs can be well mapped onto averaged minimal polymer models for the purpose of prediction of the IDPs dimensions and dynamic relaxation times. The coarse-grained simulations are in a good agreement with the results of atomistic MD. We discuss the implications of these results for the interpretation of the recent experimental measurements, and for the further applications of mesoscopic models of FG nucleoporins and IDPs more broadly.
Keywords: SAXS; amino acid sequence; end-to-end distance; hydrodynamic interactions; intrinsically disordered proteins; radius of gyration; sequence charge decoration.
Conflict of interest statement
The authors declare no competing interest.
Figures





References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
