

Influenza A Virus M1 Protein Non-Specifically Deforms Charged Lipid Membranes and Specifically Interacts with the Raft Boundary
- PMID: 36676883
- PMCID: PMC9864314
- DOI: 10.3390/membranes13010076
Influenza A Virus M1 Protein Non-Specifically Deforms Charged Lipid Membranes and Specifically Interacts with the Raft Boundary
Abstract
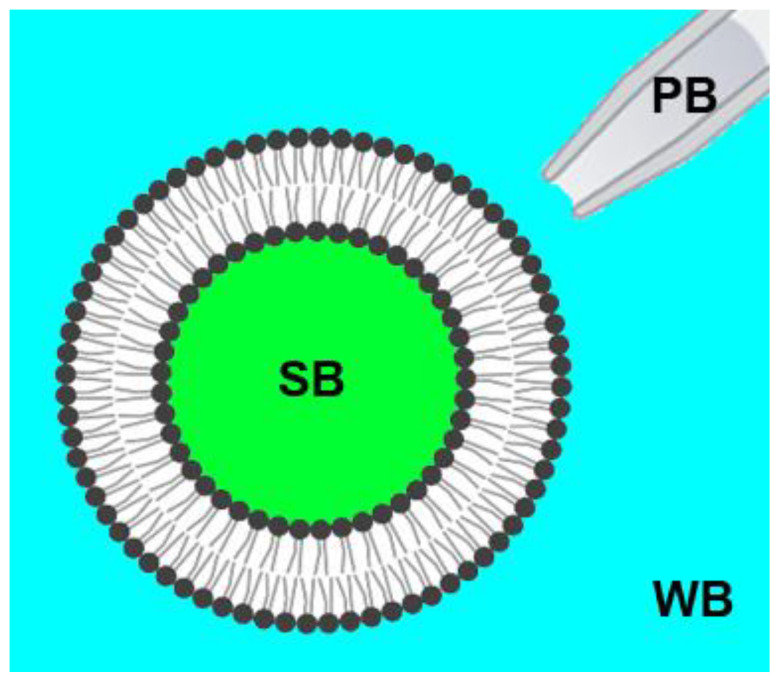
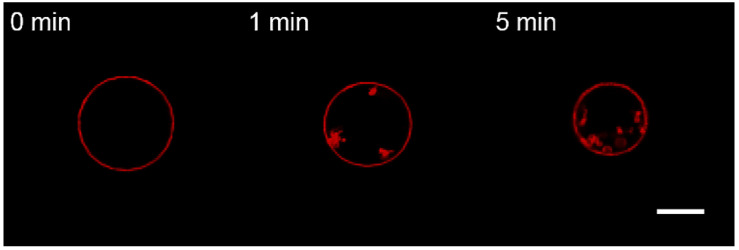
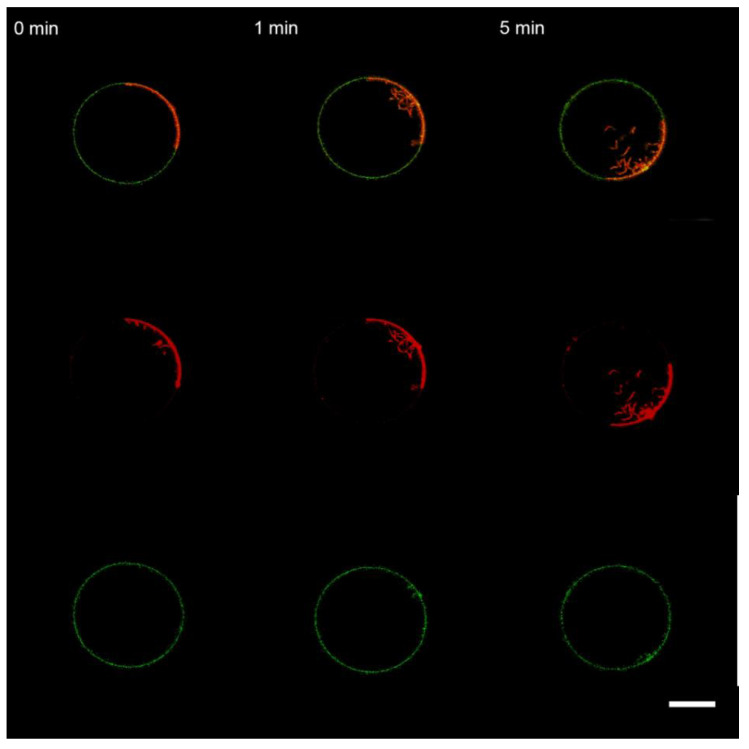
Topological rearrangements of biological membranes, such as fusion and fission, often require a sophisticated interplay between different proteins and cellular membranes. However, in the case of fusion proteins of enveloped viruses, even one molecule can execute membrane restructurings. Growing evidence indicates that matrix proteins of enveloped viruses can solely trigger the membrane bending required for another crucial step in virogenesis, the budding of progeny virions. For the case of the influenza A virus matrix protein M1, different studies report both in favor and against M1 being able to produce virus-like particles without other viral proteins. Here, we investigated the physicochemical mechanisms of M1 membrane activity on giant unilamellar vesicles of different lipid compositions using fluorescent confocal microscopy. We confirmed that M1 predominantly interacts electrostatically with the membrane, and its ability to deform the lipid bilayer is non-specific and typical for membrane-binding proteins and polypeptides. However, in the case of phase-separating membranes, M1 demonstrates a unique ability to induce macro-phase separation, probably due to the high affinity of M1's amphipathic helices to the raft boundary. Thus, we suggest that M1 is tailored to deform charged membranes with a specific activity in the case of phase-separating membranes.
Keywords: M1 matrix protein; amphipathic helices; fluorescent confocal microscopy; giant unilamellar vesicle (GUV); hemagglutinin; influenza A virus; lipid raft; lipoprotein envelope; membrane deformation; viral budding.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of this study; in the collection, analysis, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
Figures







Similar articles
-
Phosphatidylserine Lateral Organization Influences the Interaction of Influenza Virus Matrix Protein 1 with Lipid Membranes.J Virol. 2017 May 26;91(12):e00267-17. doi: 10.1128/JVI.00267-17. Print 2017 Jun 15. J Virol. 2017. PMID: 28356535 Free PMC article.
-
The Cytoplasmic Tail of Influenza A Virus Hemagglutinin and Membrane Lipid Composition Change the Mode of M1 Protein Association with the Lipid Bilayer.Membranes (Basel). 2021 Oct 10;11(10):772. doi: 10.3390/membranes11100772. Membranes (Basel). 2021. PMID: 34677538 Free PMC article.
-
Cold co-extraction of hemagglutinin and matrix M1 protein from influenza virus A by a combination of non-ionic detergents allows for visualization of the raft-like nature of the virus envelope.Arch Virol. 2008;153(10):1977-80. doi: 10.1007/s00705-008-0214-7. Epub 2008 Sep 30. Arch Virol. 2008. PMID: 18825482
-
Transport of viral proteins to the apical membranes and interaction of matrix protein with glycoproteins in the assembly of influenza viruses.Virus Res. 2001 Sep;77(1):61-9. doi: 10.1016/s0168-1702(01)00266-0. Virus Res. 2001. PMID: 11451488 Review.
-
[The concept of transmembrane asymmetry of lateral domains in biomemranes and influenza virus envelope fine structure].Mol Biol (Mosk). 2009 Jul-Aug;43(4):579-89. Mol Biol (Mosk). 2009. PMID: 19807018 Review. Russian.
Cited by
-
Odor Discrimination by Lipid Membranes.Membranes (Basel). 2023 Jan 24;13(2):151. doi: 10.3390/membranes13020151. Membranes (Basel). 2023. PMID: 36837654 Free PMC article.
-
Tailored Intranasal Albumin Caged Selegiline-α Synuclein siRNA Liposome with Improved Efficiency in Parkinson's Model.Pharmaceutics. 2025 Feb 12;17(2):243. doi: 10.3390/pharmaceutics17020243. Pharmaceutics. 2025. PMID: 40006609 Free PMC article.
-
HIV-1 Gag Polyprotein Affinity to the Lipid Membrane Is Independent of Its Surface Charge.Biomolecules. 2024 Aug 29;14(9):1086. doi: 10.3390/biom14091086. Biomolecules. 2024. PMID: 39334852 Free PMC article.
-
Human Immunodeficiency Virus Type 1 Gag Polyprotein Modulates Membrane Physical Properties like a Surfactant: Potential Implications for Virus Assembly.ACS Infect Dis. 2024 Aug 9;10(8):2870-2885. doi: 10.1021/acsinfecdis.4c00251. Epub 2024 Jun 25. ACS Infect Dis. 2024. PMID: 38917054 Free PMC article.
-
The Effects of Viral Structural Proteins on Acidic Phospholipids in Host Membranes.Viruses. 2024 Oct 31;16(11):1714. doi: 10.3390/v16111714. Viruses. 2024. PMID: 39599829 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
