

Neurobiological Basis of Aversion-Resistant Ethanol Seeking in C. elegans
- PMID: 36676987
- PMCID: PMC9861758
- DOI: 10.3390/metabo13010062
Neurobiological Basis of Aversion-Resistant Ethanol Seeking in C. elegans
Abstract
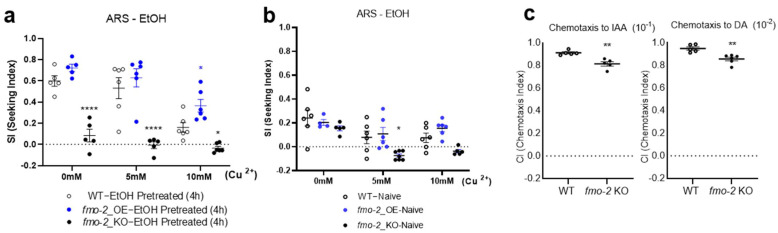
Persistent alcohol seeking despite the risk of aversive consequences is a crucial characteristic of alcohol use disorders (AUDs). Therefore, an improved understanding of the molecular basis of alcohol seeking despite aversive stimuli or punishment in animal models is an important strategy to understand the mechanism that underpins the pathology of AUDs. Aversion-resistant seeking (ARS) is characterized by disruption in control of alcohol use featured by an imbalance between the urge for alcohol and the mediation of aversive stimuli. We exploited C. elegans, a genetically tractable invertebrate, as a model to elucidate genetic components related to this behavior. We assessed the seb-3 neuropeptide system and its transcriptional regulation to progress aversion-resistant ethanol seeking at the system level. Our functional genomic approach preferentially selected molecular components thought to be involved in cholesterol metabolism, and an orthogonal test defined functional roles in ARS through behavioral elucidation. Our findings suggest that fmo-2 (flavin-containing monooxygenase-2) plays a role in the progression of aversion-resistant ethanol seeking in C. elegans.
Keywords: C. elegans; aversion-resistant seeking; ethanol preference; fmo-2; seb-3.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Neuropeptidergic regulation of compulsive ethanol seeking in C. elegans.Sci Rep. 2022 Feb 2;12(1):1804. doi: 10.1038/s41598-022-05256-1. Sci Rep. 2022. PMID: 35110557 Free PMC article.
-
Rewarding and aversive effects of ethanol in High Drinking in the Dark selectively bred mice.Addict Biol. 2015 Jan;20(1):80-90. doi: 10.1111/adb.12079. Epub 2013 Jul 30. Addict Biol. 2015. PMID: 23910826 Free PMC article.
-
Sex Differences in Ethanol Reward Seeking Under Conflict in Mice.Alcohol Clin Exp Res. 2019 Jul;43(7):1556-1566. doi: 10.1111/acer.14070. Epub 2019 May 18. Alcohol Clin Exp Res. 2019. PMID: 31034618 Free PMC article.
-
Phenotypic and genotypic characterization of the Indiana University rat lines selectively bred for high and low alcohol preference.Behav Genet. 2002 Sep;32(5):363-88. doi: 10.1023/a:1020266306135. Behav Genet. 2002. PMID: 12405517 Review.
-
Behavioral and neurochemical studies in distinct animal models of ethanol's motivational effects.Curr Drug Abuse Rev. 2010 Dec;3(4):205-21. doi: 10.2174/1874473711003040205. Curr Drug Abuse Rev. 2010. PMID: 21208169 Review.
Cited by
-
JNK Signaling Positively Regulates Acute Ethanol Tolerance in C. elegans.Int J Mol Sci. 2024 Jun 10;25(12):6398. doi: 10.3390/ijms25126398. Int J Mol Sci. 2024. PMID: 38928105 Free PMC article.
-
Mitochondria in the Spotlight: C. elegans as a Model Organism to Evaluate Xenobiotic-Induced Dysfunction.Cells. 2023 Aug 22;12(17):2124. doi: 10.3390/cells12172124. Cells. 2023. PMID: 37681856 Free PMC article. Review.
References
-
- World Health O. Global Status Report on Alcohol and Health 2018. World Health Organization; Geneva, Switzerland: 2018.
-
- Esser M.B., Sherk A., Liu Y., Naimi T.S., Stockwell T., Stahre M., Kanny D., Landen M., Saitz R., Brewer R.D. Deaths and Years of Potential Life Lost from Excessive Alcohol Use — United States, 2011–2015. MMWR Morb. Mortal. Wkly. Rep. 2020;69:1428–1433. doi: 10.15585/mmwr.mm6939a6. - DOI - PMC - PubMed
-
- Rehm J., Baliunas D., Borges G.L., Graham K., Irving H., Kehoe T., Parry C.D., Patra J., Popova S., Poznyak V., et al. The relation between different dimensions of alcohol consumption and burden of disease: An overview. Addiction. 2010;105:817–843. doi: 10.1111/j.1360-0443.2010.02899.x. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
