

Recent Insights into Plant miRNA Biogenesis: Multiple Layers of miRNA Level Regulation
- PMID: 36679055
- PMCID: PMC9864873
- DOI: 10.3390/plants12020342
Recent Insights into Plant miRNA Biogenesis: Multiple Layers of miRNA Level Regulation
Abstract
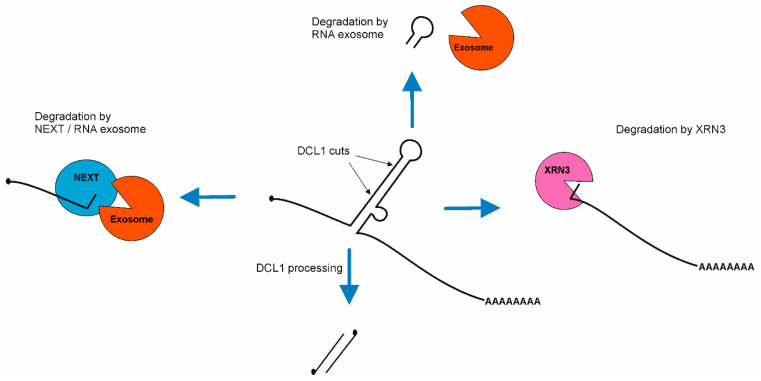
MicroRNAs are small RNAs, 20-22 nt long, the main role of which is to downregulate gene expression at the level of mRNAs. MiRNAs are fundamental regulators of plant growth and development in response to internal signals as well as in response to abiotic and biotic factors. Therefore, the deficiency or excess of individual miRNAs is detrimental to particular aspects of a plant's life. In consequence, the miRNA levels must be appropriately adjusted. To obtain proper expression of each miRNA, their biogenesis is controlled at multiple regulatory layers. Here, we addressed processes discovered to influence miRNA steady-state levels, such as MIR transcription, co-transcriptional pri-miRNA processing (including splicing, polyadenylation, microprocessor assembly and activity) and miRNA-encoded peptides synthesis. MiRNA stability, RISC formation and miRNA export out of the nucleus and out of the plant cell also define the levels of miRNAs in various plant tissues. Moreover, we show the evolutionary conservation of miRNA biogenesis core proteins across the plant kingdom.
Keywords: Arabidopsis; PTGS; core microprocessor evolutionary conservation; miRNA biogenesis; microRNA; microprocessor; plants; pri-miRNA degradation; regulation of miRNA level.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Publication types
LinkOut - more resources
Full Text Sources
Research Materials
