

Iron Nutrition in Plants: Towards a New Paradigm?
- PMID: 36679097
- PMCID: PMC9862363
- DOI: 10.3390/plants12020384
Iron Nutrition in Plants: Towards a New Paradigm?
Abstract
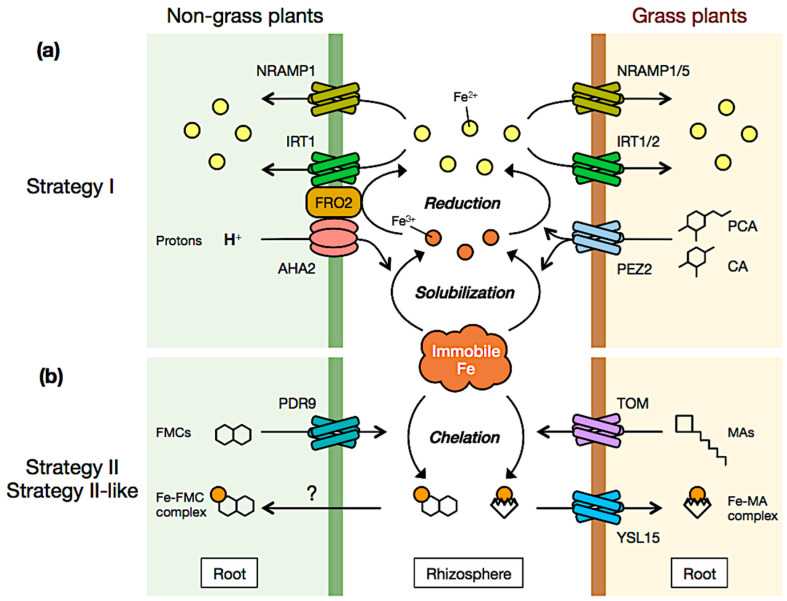
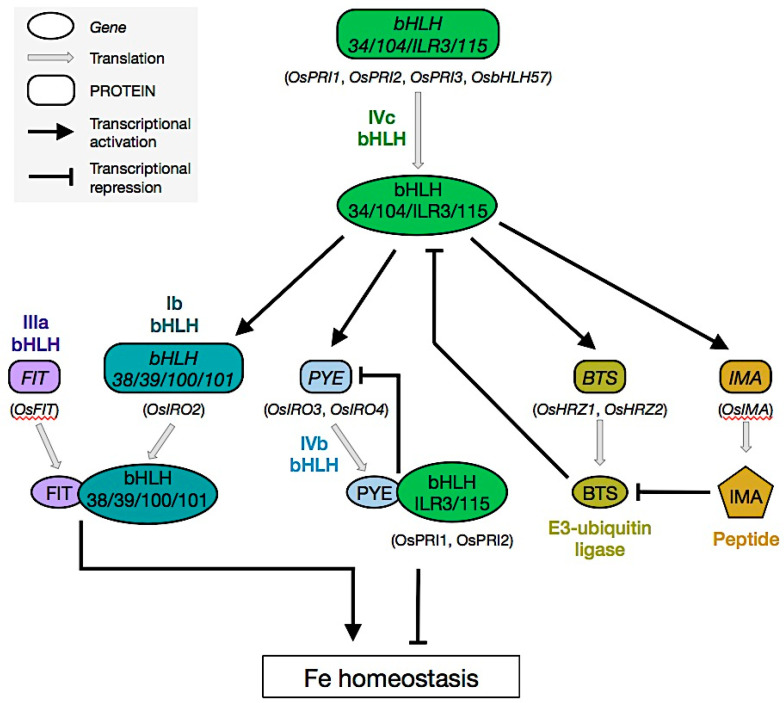
Iron (Fe) is an essential micronutrient for plant growth and development. Fe availability affects crops' productivity and the quality of their derived products and thus human nutrition. Fe is poorly available for plant use since it is mostly present in soils in the form of insoluble oxides/hydroxides, especially at neutral to alkaline pH. How plants cope with low-Fe conditions and acquire Fe from soil has been investigated for decades. Pioneering work highlighted that plants have evolved two different strategies to mine Fe from soils, the so-called Strategy I (Fe reduction strategy) and Strategy II (Fe chelation strategy). Strategy I is employed by non-grass species whereas graminaceous plants utilize Strategy II. Recently, it has emerged that these two strategies are not fully exclusive and that the mechanism used by plants for Fe uptake is directly shaped by the characteristics of the soil on which they grow (e.g., pH, oxygen concentration). In this review, recent findings on plant Fe uptake and the regulation of this process will be summarized and their impact on our understanding of plant Fe nutrition will be discussed.
Keywords: Arabidopsis; bHLH; dicots; grass; iron homeostasis; transcription factor.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the writing of the manuscript.
Figures
References
-
- Touraine B., Vignols F., Przybyla-Toscano J., Ischebeck T., Dhalleine T., Wu H.C., Magno C., Berger N., Couturier J., Dubos C., et al. Iron-sulfur protein NFU2 is required for branched-chain amino acid synthesis in Arabidopsis roots. J. Exp. Bot. 2019;70:1875–1889. doi: 10.1093/jxb/erz050. - DOI - PubMed
-
- Becker M., Asch F. Iron toxicity in rice-conditions and management concepts. J. Plant Nutr. Soil Sci. 2005;168:558–573. doi: 10.1002/jpln.200520504. - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
