

A Polydnavirus Protein Tyrosine Phosphatase Negatively Regulates the Host Phenoloxidase Pathway
- PMID: 36680096
- PMCID: PMC9866809
- DOI: 10.3390/v15010056
A Polydnavirus Protein Tyrosine Phosphatase Negatively Regulates the Host Phenoloxidase Pathway
Abstract
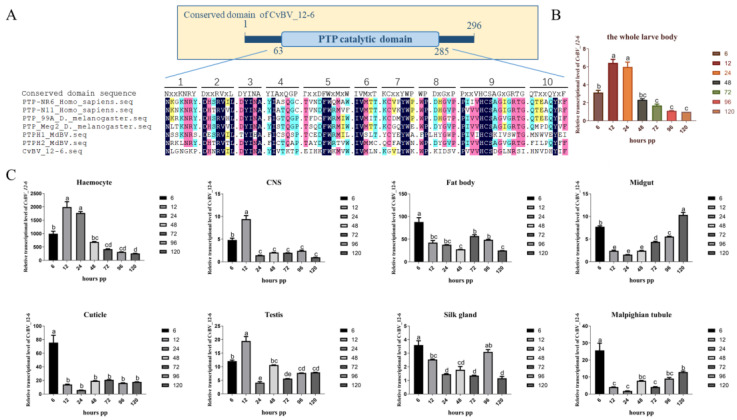
Polydnavirus (PDV) is a parasitic factor of endoparasitic wasps and contributes greatly to overcoming the immune response of parasitized hosts. Protein tyrosine phosphatases (PTPs) regulate a wide variety of biological processes at the post-transcriptional level in mammals, but knowledge of PDV PTP action during a parasitoid−host interaction is limited. In this study, we characterized a PTP gene, CvBV_12-6, derived from Cotesia vestalis bracovirus (CvBV), and explored its possible regulatory role in the immune response of the host Plutella xylostella. Our results from qPCR show that CvBV_12-6 was highly expressed in hemocytes at an early stage of parasitization. To explore CvBV_12-6 function, we specifically expressed CvBV_12-6 in Drosophila melanogaster hemocytes. The results show that Hml-Gal4 > CvBV_12-6 suppressed the phenoloxidase activity of hemolymph in D. melanogaster, but exerted no effect on the total count or the viability of the hemocytes. In addition, the Hml-Gal4 > CvBV_12-6 flies exhibited decreased antibacterial abilities against Staphylococcus aureus. Similarly, we found that CvBV_12-6 significantly suppressed the melanization of the host P. xylostella 24 h post parasitization and reduced the viability, but not the number, of hemocytes. In conclusion, CvBV_12-6 negatively regulated both cellular and humoral immunity in P. xylostella, and the related molecular mechanism may be universal to insects.
Keywords: phenoloxidase; polydnavirus; protein tyrosine phosphatase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
