

Glucocorticoids coordinate the bladder peripheral clock and diurnal micturition pattern in mice
- PMID: 36681730
- PMCID: PMC9867708
- DOI: 10.1038/s42003-023-04464-3
Glucocorticoids coordinate the bladder peripheral clock and diurnal micturition pattern in mice
Abstract
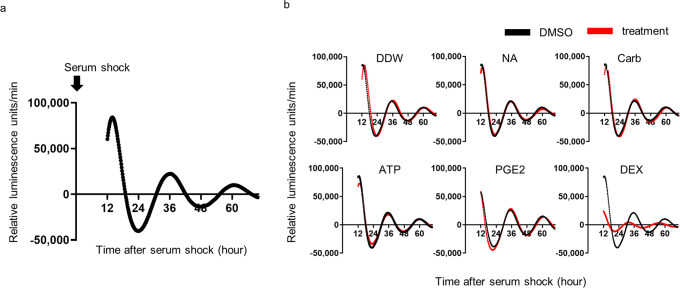
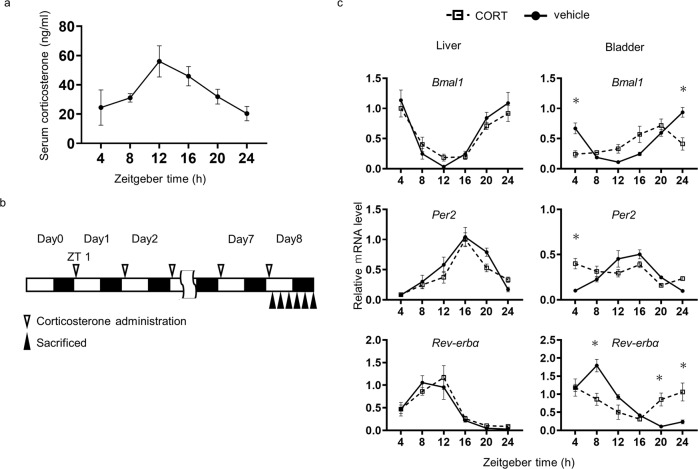
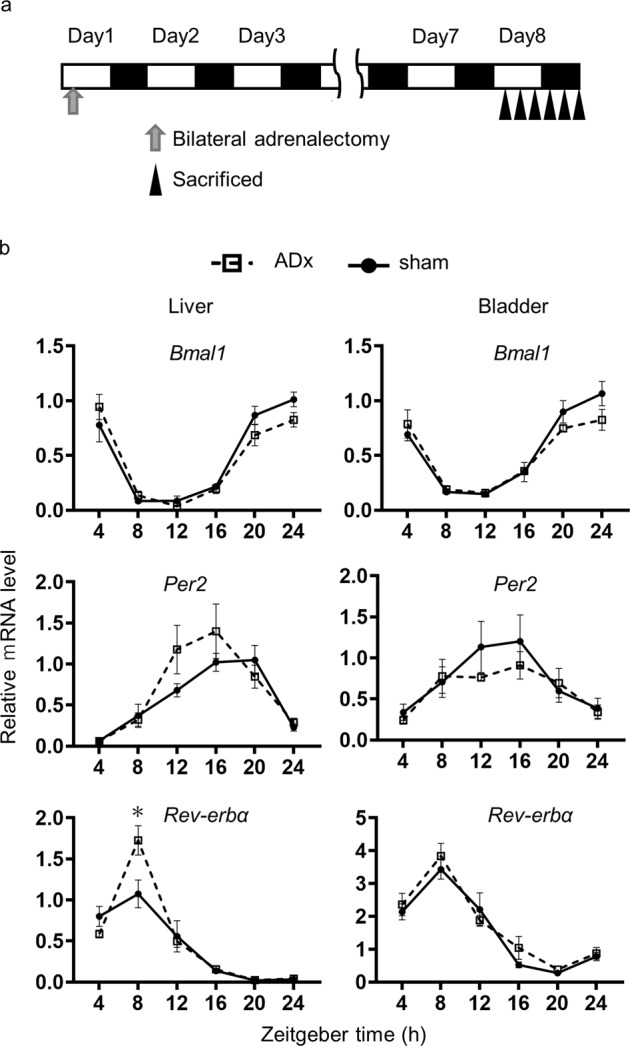
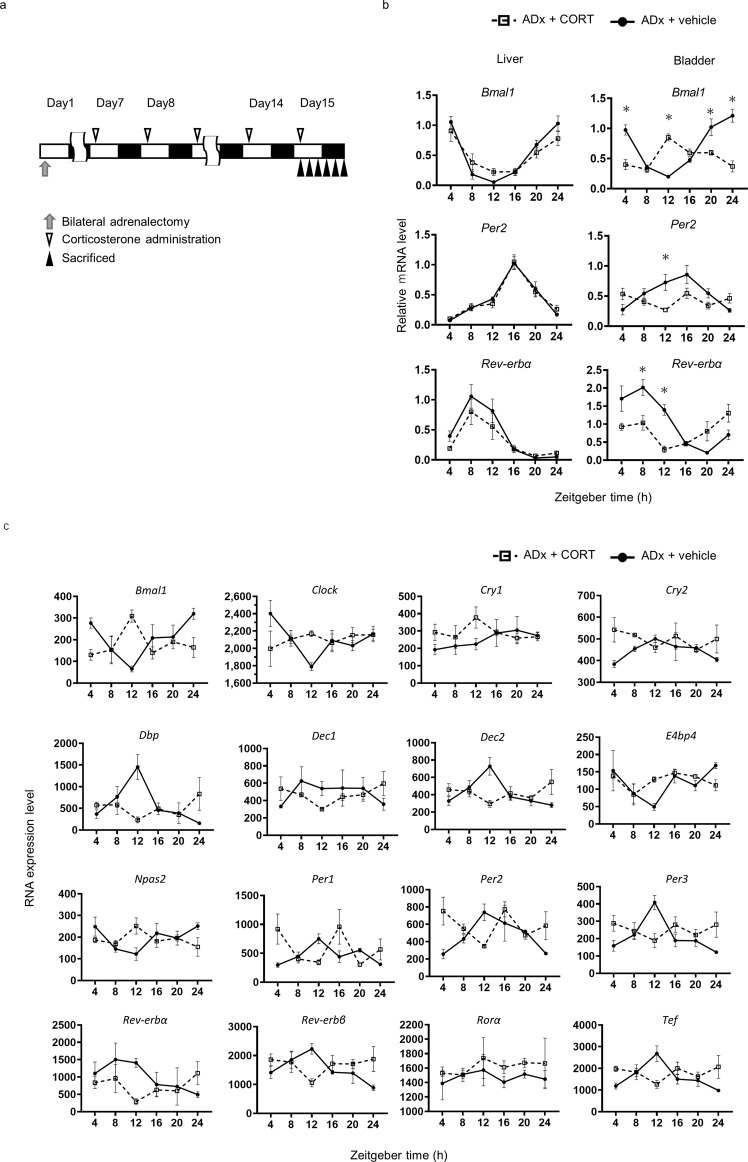
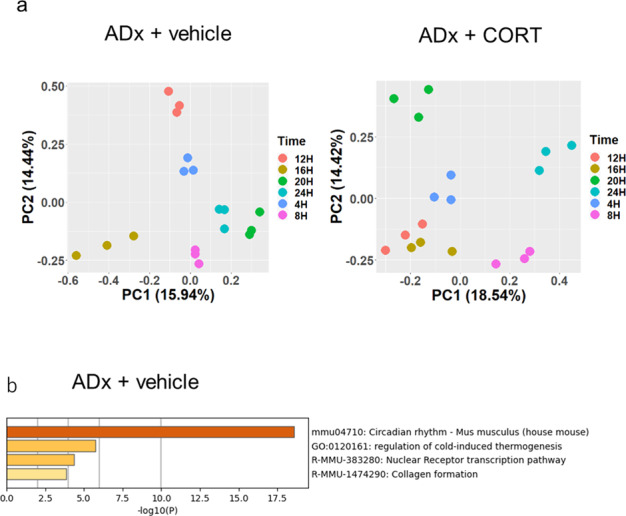
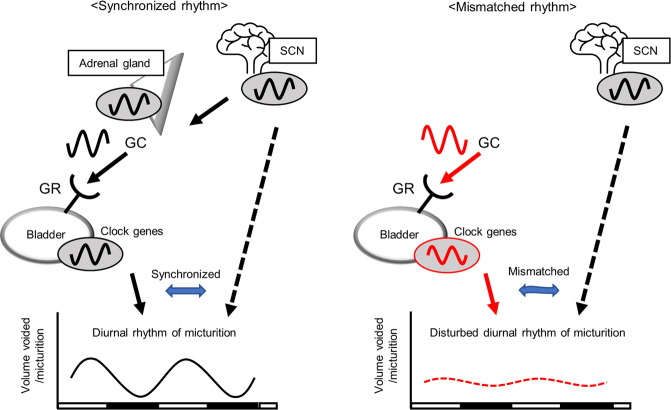
Peripheral clocks function to regulate each organ and are synchronized though various molecular and behavioral signals. However, signals that entrain the bladder clock remain elusive. Here, we show that glucocorticoids are a key cue for the bladder clock in vitro and in vivo. A pBmal1-dLuc human urothelial cell-line showed significant shifts in gene expression after cortisol treatment. In vivo, rhythmic bladder clock gene expression was unchanged by bilateral adrenalectomy but shifted 4 h forward by corticosterone administration at the inactive phase. Moreover, the bladder clock shifted 8-12 h in mice that underwent both bilateral adrenalectomy and corticosterone administration at the inactive phase. These mice showed decreases in the diurnal rhythm of volume voided per micturition, while maintaining diurnal activity rhythms. These results indicate that the diurnal rhythm of glucocorticoid signaling is a zeitgeber that overcomes other bladder clock entrainment factors and coordinates the diurnal rhythm of volume voided per micturition.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Chronobiology of micturition: putative role of the circadian clock.J Urol. 2013 Sep;190(3):843-9. doi: 10.1016/j.juro.2013.02.024. Epub 2013 Feb 18. J Urol. 2013. PMID: 23429068 Review.
-
Feeding and adrenal entrainment stimuli are both necessary for normal circadian oscillation of peripheral clocks in mice housed under different photoperiods.Chronobiol Int. 2015 Mar;32(2):195-210. doi: 10.3109/07420528.2014.962655. Epub 2014 Oct 6. Chronobiol Int. 2015. PMID: 25286135
-
Phase shifts in circadian peripheral clocks caused by exercise are dependent on the feeding schedule in PER2::LUC mice.Chronobiol Int. 2016;33(7):849-62. doi: 10.3109/07420528.2016.1171775. Epub 2016 Apr 28. Chronobiol Int. 2016. PMID: 27123825
-
Altered entrainment to the day/night cycle attenuates the daily rise in circulating corticosterone in the mouse.PLoS One. 2014 Nov 3;9(11):e111944. doi: 10.1371/journal.pone.0111944. eCollection 2014. PLoS One. 2014. PMID: 25365210 Free PMC article.
-
Temperature effect on entrainment, phase shifting, and amplitude of circadian clocks and its molecular bases.Chronobiol Int. 2002 Sep;19(5):807-64. doi: 10.1081/cbi-120014569. Chronobiol Int. 2002. PMID: 12405549 Review.
Cited by
-
Disruption of circadian rhythm as a potential pathogenesis of nocturia.Nat Rev Urol. 2025 May;22(5):276-293. doi: 10.1038/s41585-024-00961-0. Epub 2024 Nov 14. Nat Rev Urol. 2025. PMID: 39543359 Free PMC article. Review.
-
Exploratory metabolomic analysis for characterizing the metabolic profile of the urinary bladder under estrogen deprivation.Front Endocrinol (Lausanne). 2024 May 31;15:1384115. doi: 10.3389/fendo.2024.1384115. eCollection 2024. Front Endocrinol (Lausanne). 2024. PMID: 38883607 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
