

Single-cell transcriptomics reveal extracellular vesicles secretion with a cardiomyocyte proteostasis signature during pathological remodeling
- PMID: 36681760
- PMCID: PMC9867722
- DOI: 10.1038/s42003-022-04402-9
Single-cell transcriptomics reveal extracellular vesicles secretion with a cardiomyocyte proteostasis signature during pathological remodeling
Erratum in
-
Author Correction: Single-cell transcriptomics reveal extracellular vesicles secretion with a cardiomyocyte proteostasis signature during pathological remodeling.Commun Biol. 2024 May 23;7(1):624. doi: 10.1038/s42003-024-06319-x. Commun Biol. 2024. PMID: 38783104 Free PMC article. No abstract available.
Abstract
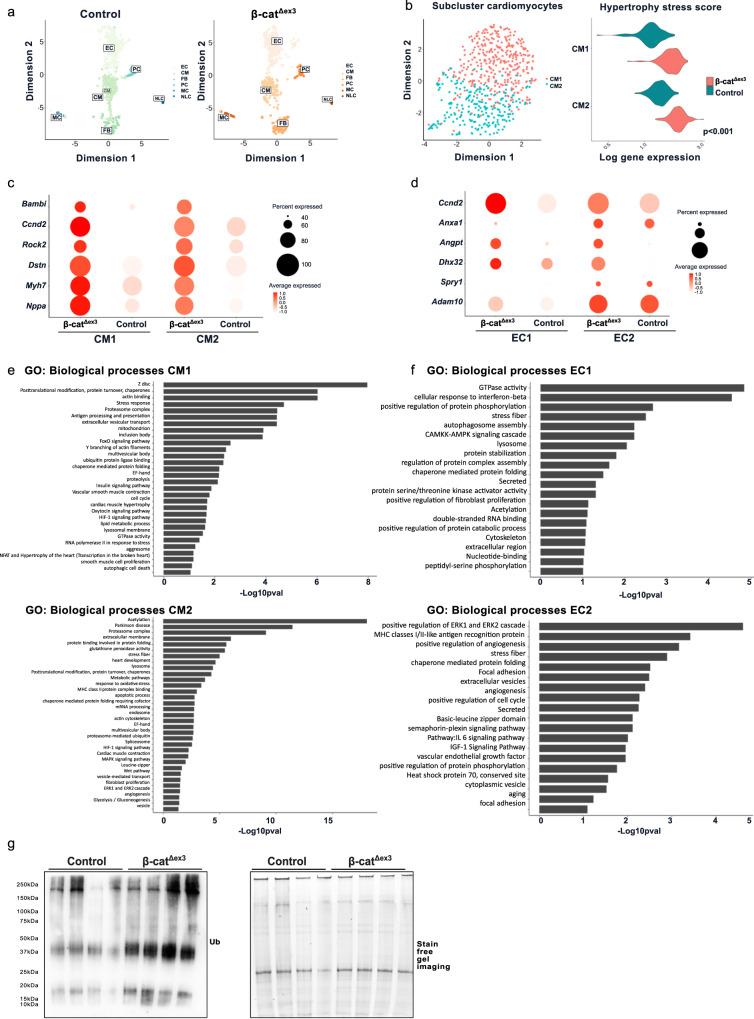
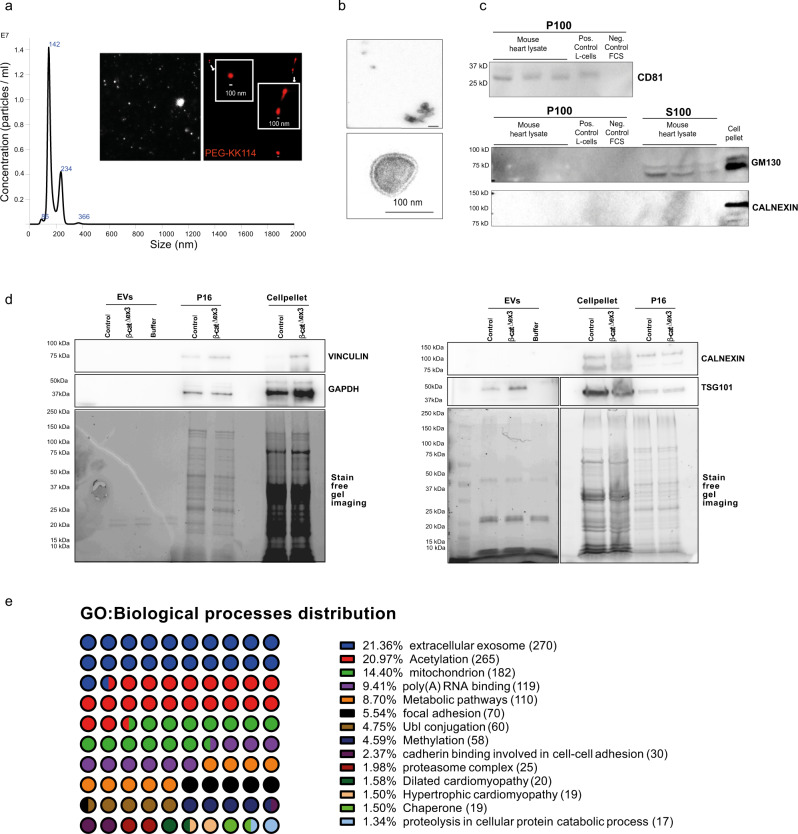
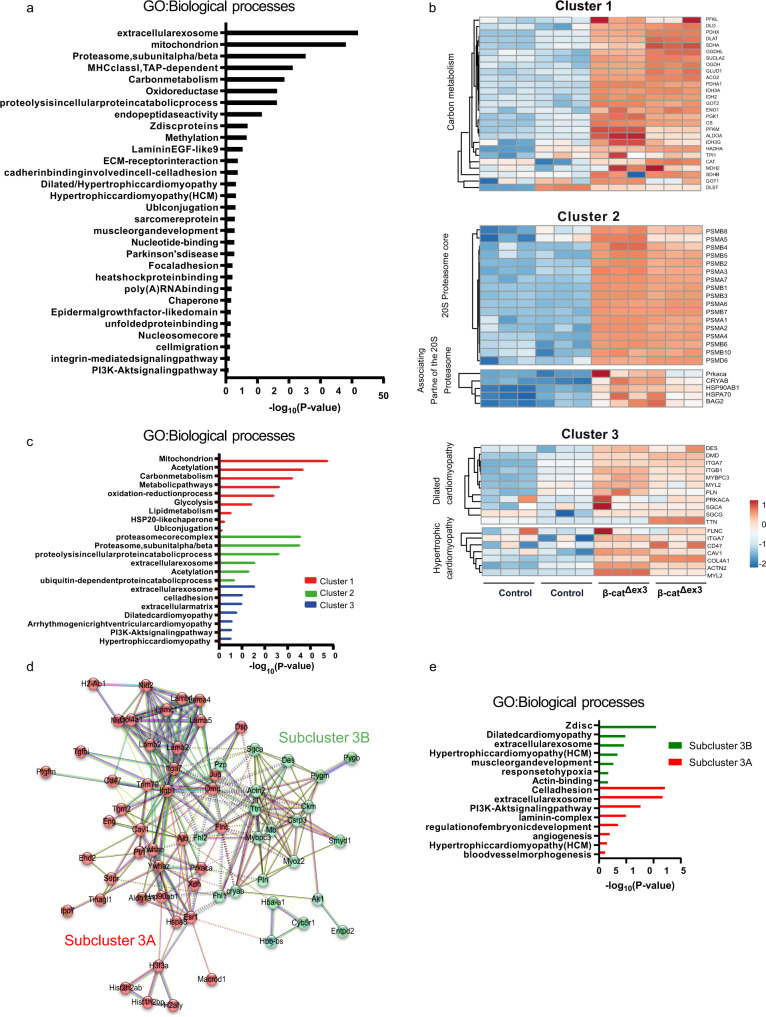
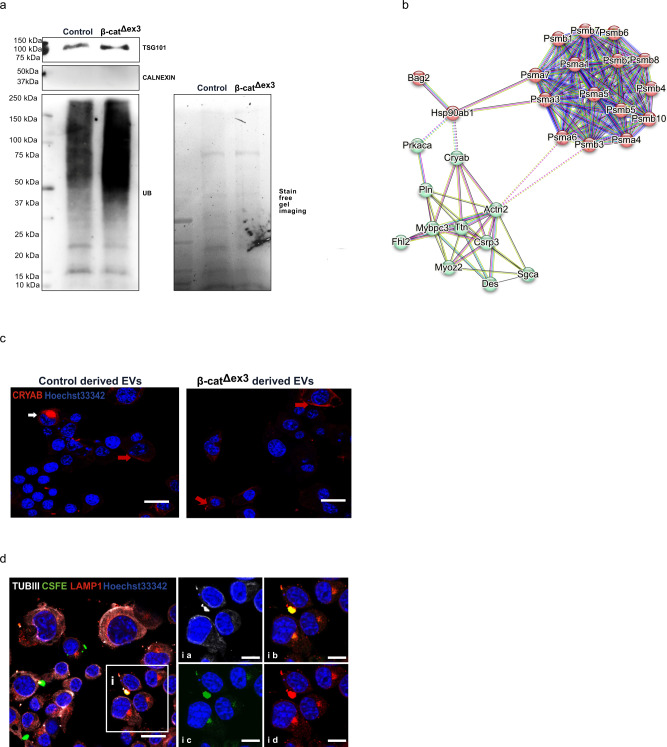
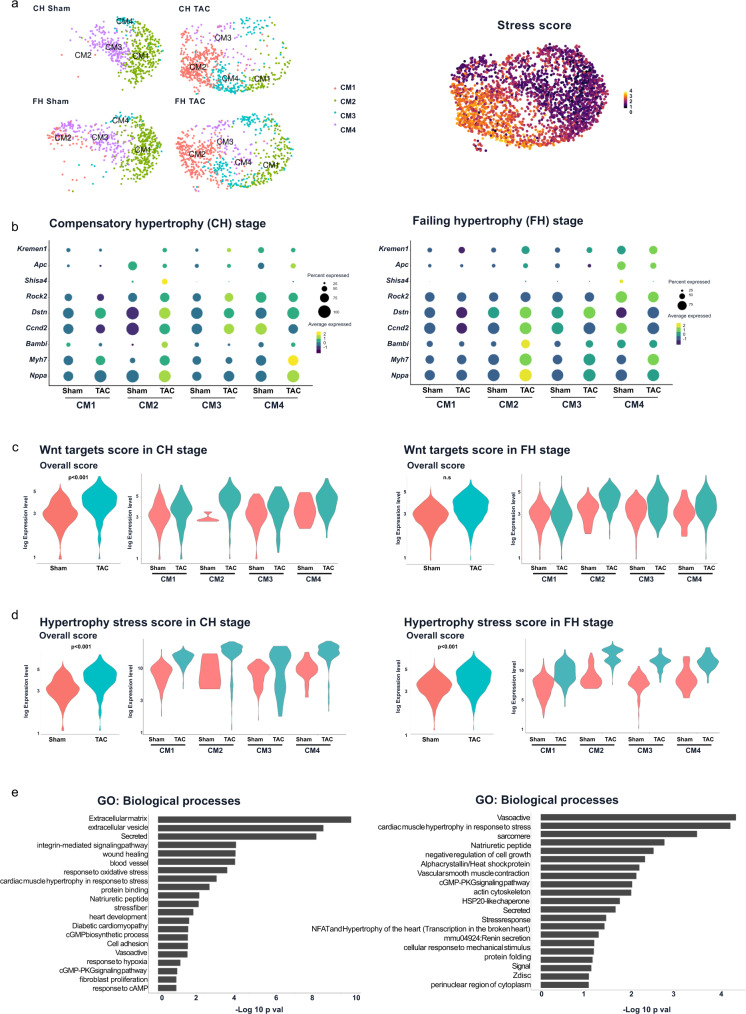
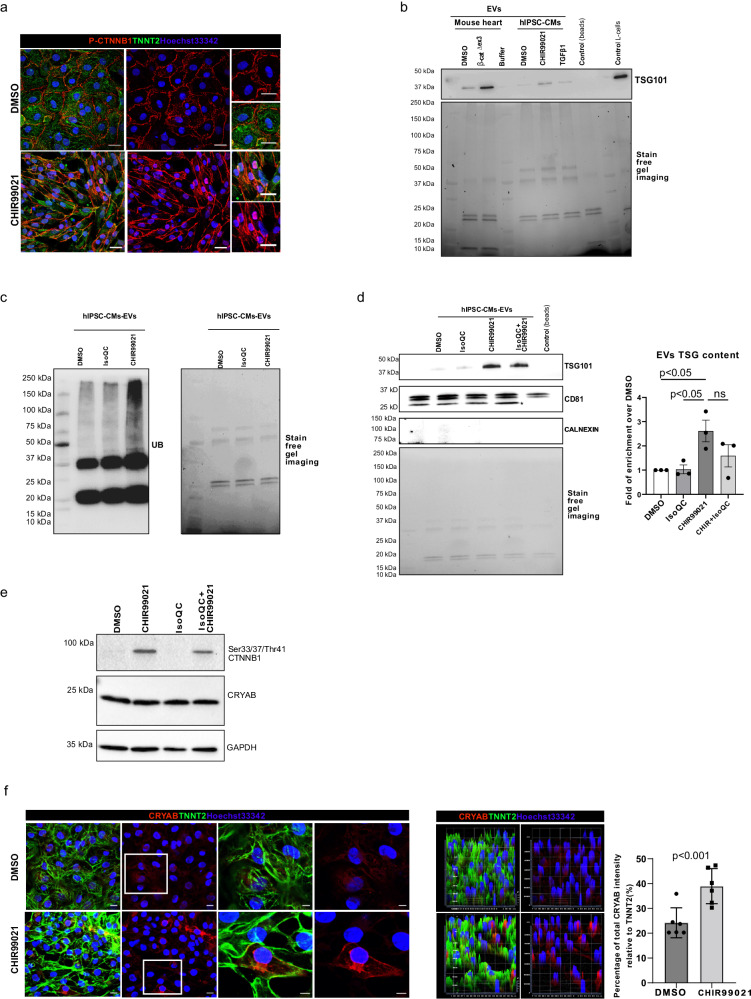
Aberrant Wnt activation has been reported in failing cardiomyocytes. Here we present single cell transcriptome profiling of hearts with inducible cardiomyocyte-specific Wnt activation (β-catΔex3) as well as with compensatory and failing hypertrophic remodeling. We show that functional enrichment analysis points to an involvement of extracellular vesicles (EVs) related processes in hearts of β-catΔex3 mice. A proteomic analysis of in vivo cardiac derived EVs from β-catΔex3 hearts has identified differentially enriched proteins involving 20 S proteasome constitutes, protein quality control (PQC), chaperones and associated cardiac proteins including α-Crystallin B (CRYAB) and sarcomeric components. The hypertrophic model confirms that cardiomyocytes reacted with an acute early transcriptional upregulation of exosome biogenesis processes and chaperones transcripts including CRYAB, which is ameliorated in advanced remodeling. Finally, human induced pluripotent stem cells (iPSC)-derived cardiomyocytes subjected to pharmacological Wnt activation recapitulated the increased expression of exosomal markers, CRYAB accumulation and increased PQC signaling. These findings reveal that secretion of EVs with a proteostasis signature contributes to early patho-physiological adaptation of cardiomyocytes, which may serve as a read-out of disease progression and can be used for monitoring cellular remodeling in vivo with a possible diagnostic and prognostic role in the future.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
