

Metabolic regulation of stress erythropoiesis, outstanding questions, and possible paradigms
- PMID: 36685181
- PMCID: PMC9849390
- DOI: 10.3389/fphys.2022.1063294
Metabolic regulation of stress erythropoiesis, outstanding questions, and possible paradigms
Abstract
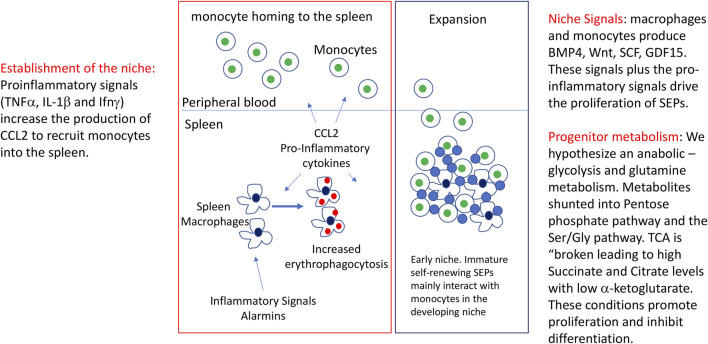
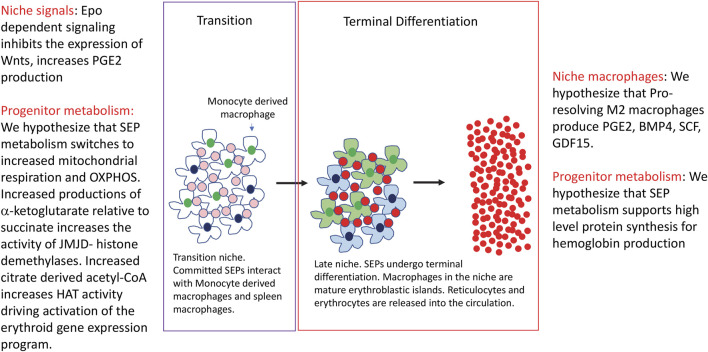
Steady state erythropoiesis produces new erythrocytes at a constant rate to replace the senescent cells that are removed by macrophages in the liver and spleen. However, infection and tissue damage disrupt the production of erythrocytes by steady state erythropoiesis. During these times, stress erythropoiesis is induced to compensate for the loss of erythroid output. The strategy of stress erythropoiesis is different than steady state erythropoiesis. Stress erythropoiesis generates a wave of new erythrocytes to maintain homeostasis until steady state conditions are resumed. Stress erythropoiesis relies on the rapid proliferation of immature progenitor cells that do not differentiate until the increase in serum Erythropoietin (Epo) promotes the transition to committed progenitors that enables their synchronous differentiation. Emerging evidence has revealed a central role for cell metabolism in regulating the proliferation and differentiation of stress erythroid progenitors. During the initial expansion stage, the immature progenitors are supported by extensive metabolic changes which are designed to direct the use of glucose and glutamine to increase the biosynthesis of macromolecules necessary for cell growth and division. At the same time, these metabolic changes act to suppress the expression of genes involved in erythroid differentiation. In the subsequent transition stage, changes in niche signals alter progenitor metabolism which in turn removes the inhibition of erythroid differentiation generating a bolus of new erythrocytes to alleviate anemia. This review summarizes what is known about the metabolic regulation of stress erythropoiesis and discusses potential mechanisms for metabolic regulation of proliferation and differentiation.
Keywords: anabolic metabolism; epigenetic regulation; glycolysis; stress erythropoiesis; tissue regeneration.
Copyright © 2023 Ruan and Paulson.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Gdf15 regulates murine stress erythroid progenitor proliferation and the development of the stress erythropoiesis niche.Blood Adv. 2019 Jul 23;3(14):2205-2217. doi: 10.1182/bloodadvances.2019000375. Blood Adv. 2019. PMID: 31324641 Free PMC article.
-
Yap1 promotes proliferation of transiently amplifying stress erythroid progenitors during erythroid regeneration.Exp Hematol. 2019 Dec;80:42-54.e4. doi: 10.1016/j.exphem.2019.11.002. Epub 2019 Nov 19. Exp Hematol. 2019. PMID: 31756359 Free PMC article.
-
Epo receptor signaling in macrophages alters the splenic niche to promote erythroid differentiation.Blood. 2020 Jul 9;136(2):235-246. doi: 10.1182/blood.2019003480. Blood. 2020. PMID: 32350523 Free PMC article.
-
Stress erythropoiesis: definitions and models for its study.Exp Hematol. 2020 Sep;89:43-54.e2. doi: 10.1016/j.exphem.2020.07.011. Epub 2020 Aug 2. Exp Hematol. 2020. PMID: 32750404 Free PMC article. Review.
-
Stress Erythropoiesis is a Key Inflammatory Response.Cells. 2020 Mar 6;9(3):634. doi: 10.3390/cells9030634. Cells. 2020. PMID: 32155728 Free PMC article. Review.
Cited by
-
Behavioral and metabolic effects of escapable electric foot shock stress in male mice.BMC Res Notes. 2025 Mar 25;18(1):127. doi: 10.1186/s13104-025-07189-0. BMC Res Notes. 2025. PMID: 40133962 Free PMC article.
-
Phenotypic Alterations in Erythroid Nucleated Cells of Spleen and Bone Marrow in Acute Hypoxia.Cells. 2023 Dec 10;12(24):2810. doi: 10.3390/cells12242810. Cells. 2023. PMID: 38132130 Free PMC article.
-
Metabolic regulation of erythrocyte development and disorders.Exp Hematol. 2024 Mar;131:104153. doi: 10.1016/j.exphem.2024.104153. Epub 2024 Jan 17. Exp Hematol. 2024. PMID: 38237718 Free PMC article.
-
Defining Candidate Imprinted loci in Bos taurus.Genes (Basel). 2023 May 2;14(5):1036. doi: 10.3390/genes14051036. Genes (Basel). 2023. PMID: 37239396 Free PMC article.
-
TFPI from erythroblasts drives heme production in central macrophages promoting erythropoiesis in polycythemia.Nat Commun. 2024 May 10;15(1):3976. doi: 10.1038/s41467-024-48328-8. Nat Commun. 2024. PMID: 38729948 Free PMC article.
References
-
- Akilesh H. M., Buechler M. B., Duggan J. M., Hahn W. O., Matta B., Sun X., et al. (2019). Chronic TLR7 and TLR9 signaling drives anemia via differentiation of specialized hemophagocytes. Science 363 (6423), eaao5213. Epub 2019/01/12PubMed PMID: 30630901; PMCID: PMC6413693. 10.1126/science.aao5213 - DOI - PMC - PubMed
-
- Arezes J., Foy N., McHugh K., Sawant A., Quinkert D., Terraube V., et al. (2018). Erythroferrone inhibits the induction of hepcidin by BMP6. Blood 132 (14), 1473–1477. Epub 2018/08/12PubMed PMID: 30097509; PMCID: PMC6238155 funding from Pfizer. N.F., A.S., V.T., A.B., M.T., E.R.L., O.C., M.L., and R.J. are Pfizer employees. N.F., O.C., R.J., J.A., K.M., S.J.D., and H.D. are named inventors on a patent application currently under evaluation. The remaining authors declare no competing financial interests. 10.1182/blood-2018-06-857995 - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials