

ADAR1 Non-Editing Function in Macrophage Activation and Abdominal Aortic Aneurysm
- PMID: 36688311
- PMCID: PMC10316962
- DOI: 10.1161/CIRCRESAHA.122.321722
ADAR1 Non-Editing Function in Macrophage Activation and Abdominal Aortic Aneurysm
Abstract
Background: Macrophage activation plays a critical role in abdominal aortic aneurysm (AAA) development. However, molecular mechanisms controlling macrophage activation and vascular inflammation in AAA remain largely unknown. The objective of the study was to identify novel mechanisms underlying adenosine deaminase acting on RNA (ADAR1) function in macrophage activation and AAA formation.
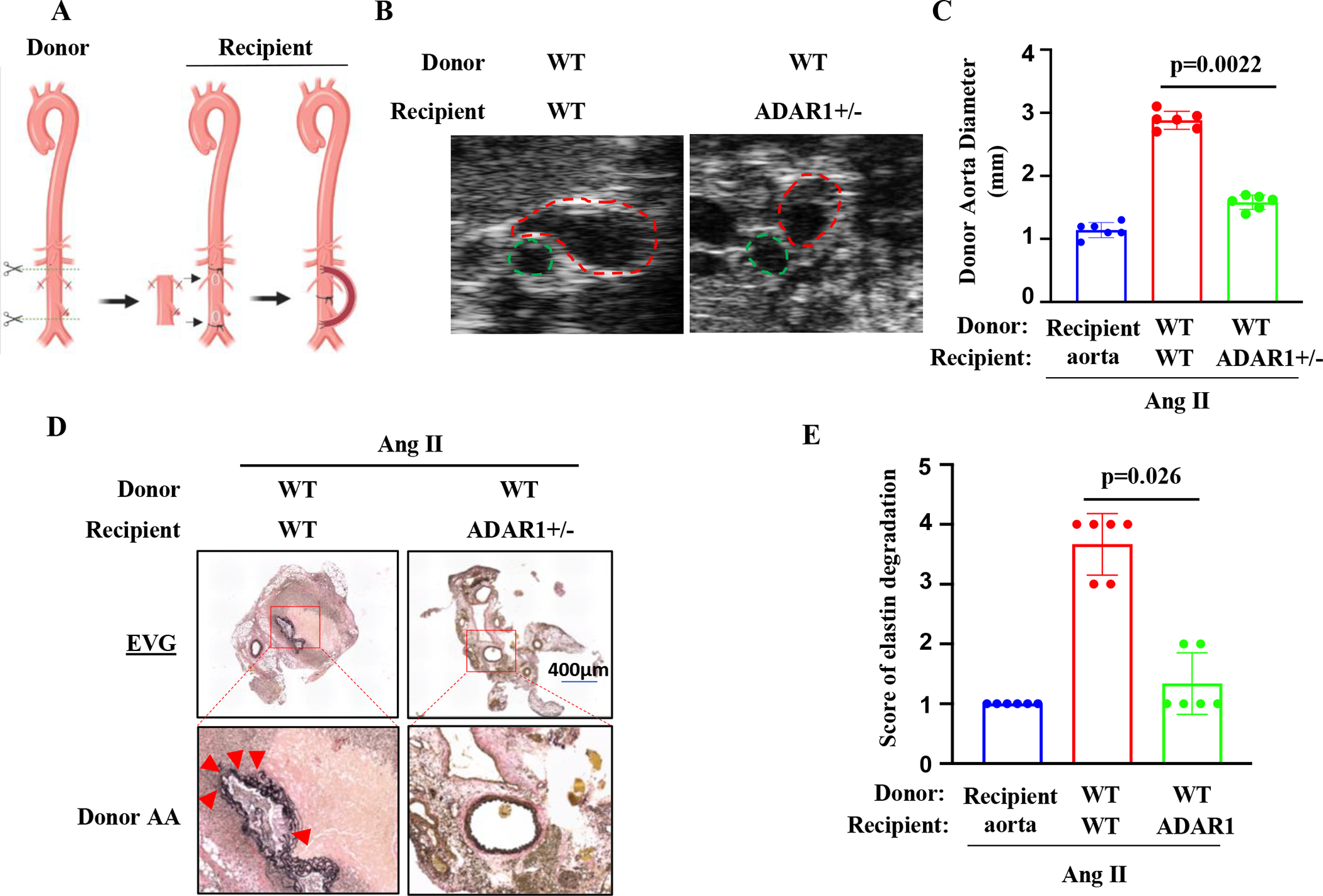
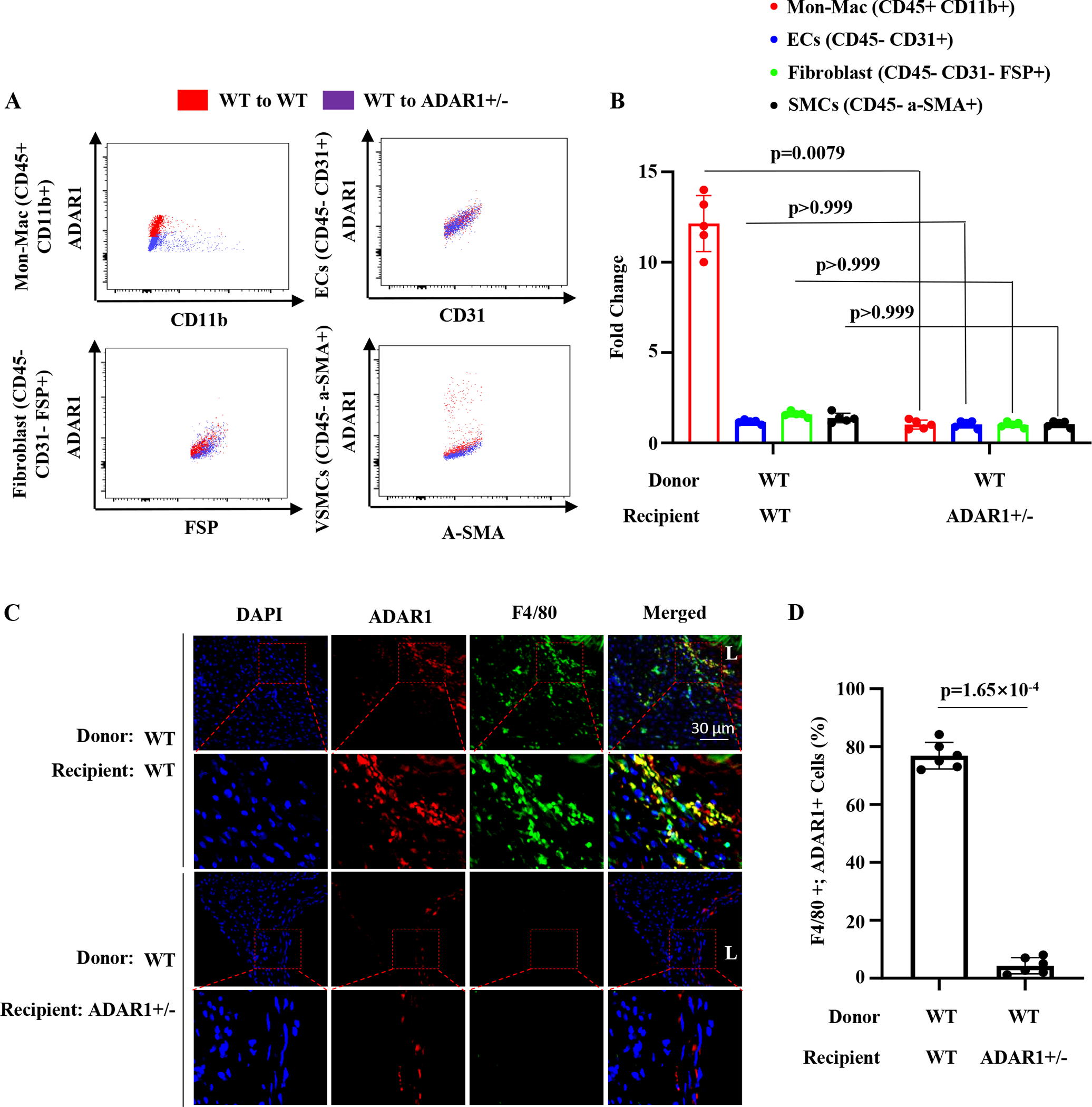
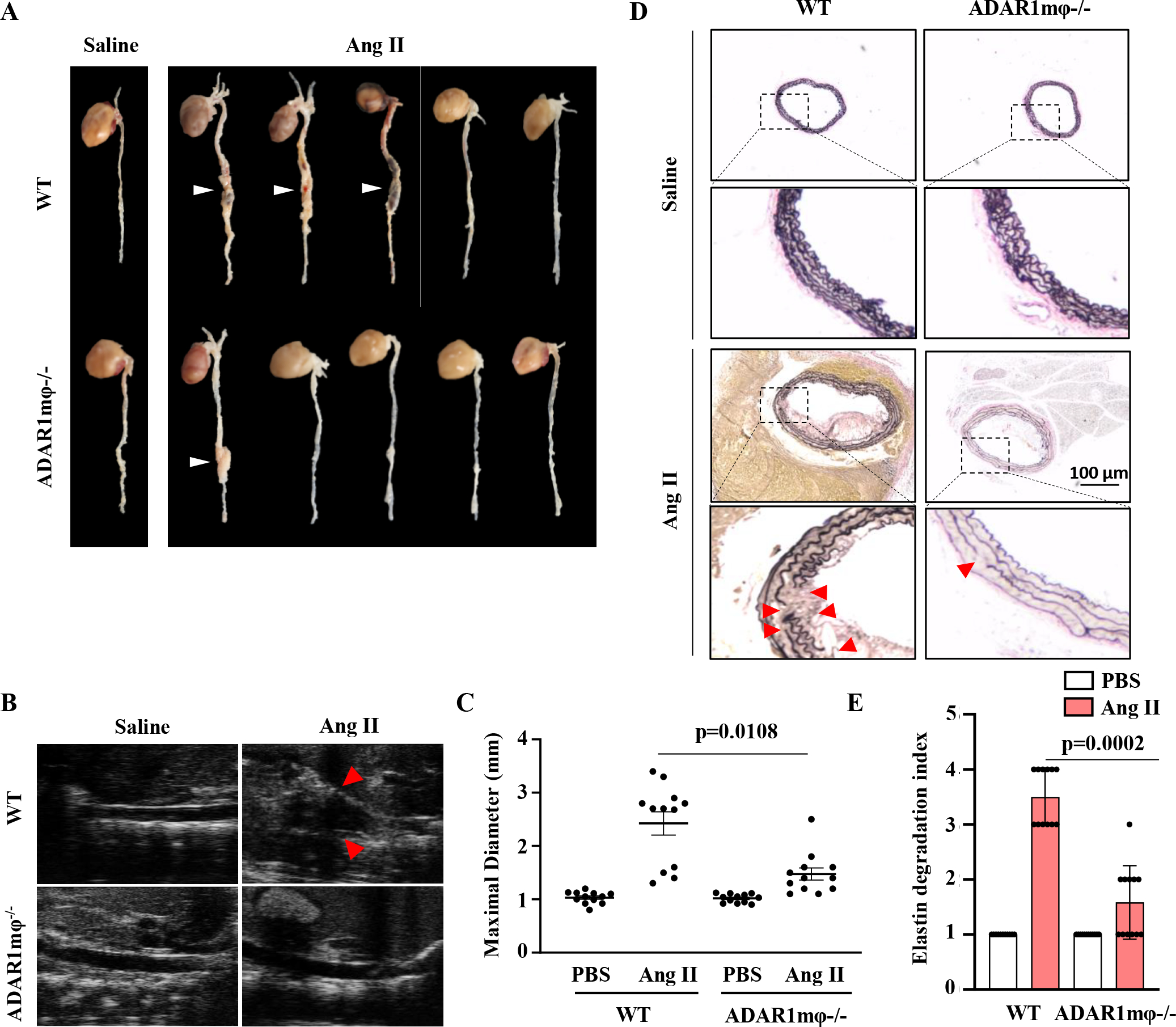
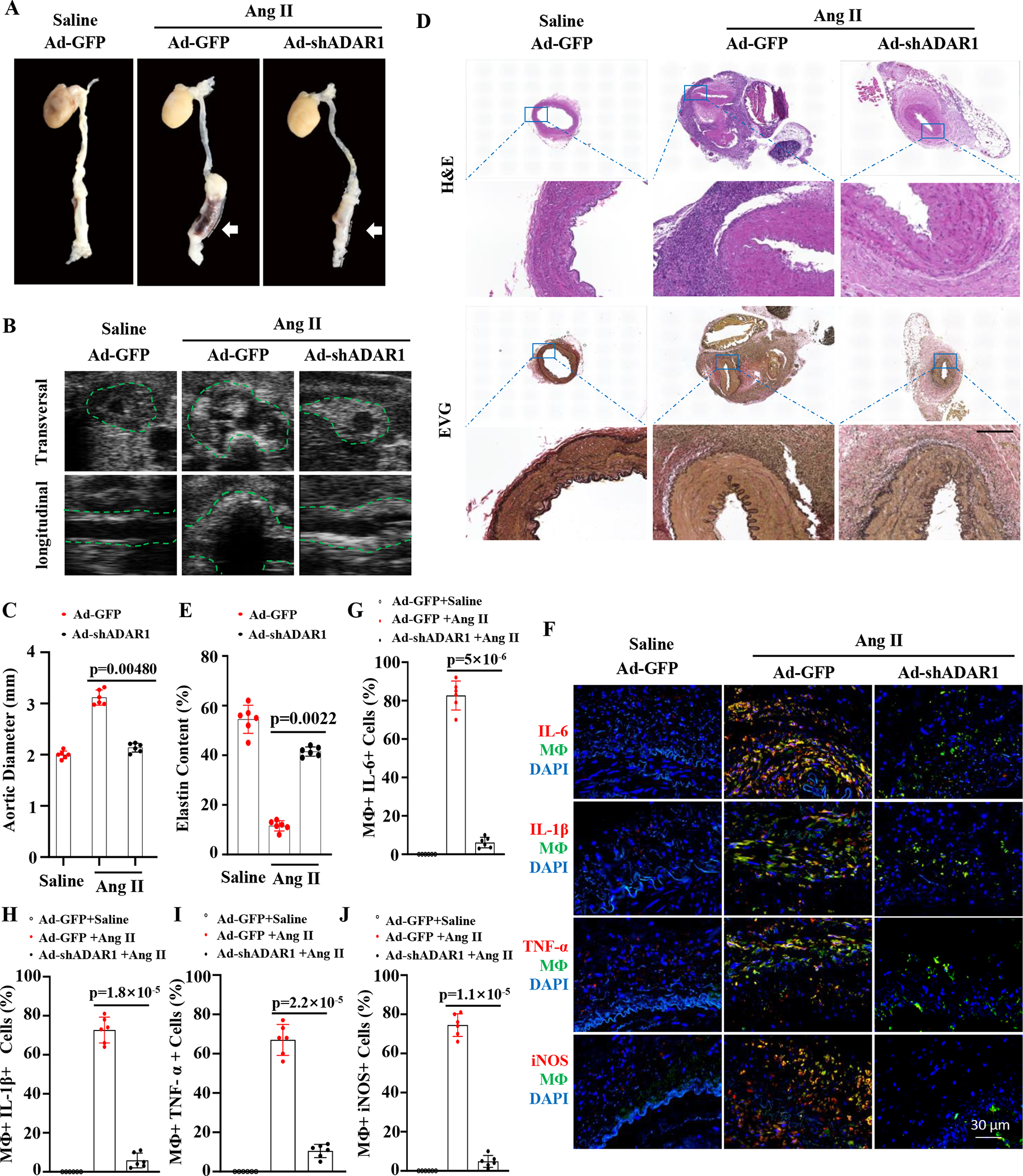
Methods: Aortic transplantation was conducted to determine the importance of nonvascular ADAR1 in AAA development/dissection. Ang II (Angiotensin II) infusion of ApoE-/- mouse model combined with macrophage-specific knockout of ADAR1 was used to study ADAR1 macrophage-specific role in AAA formation/dissection. The relevance of macrophage ADAR1 to human AAA was examined using human aneurysm specimens. Moreover, a novel humanized AAA model was established to test the role of human macrophages in aneurysm formation in human arteries.
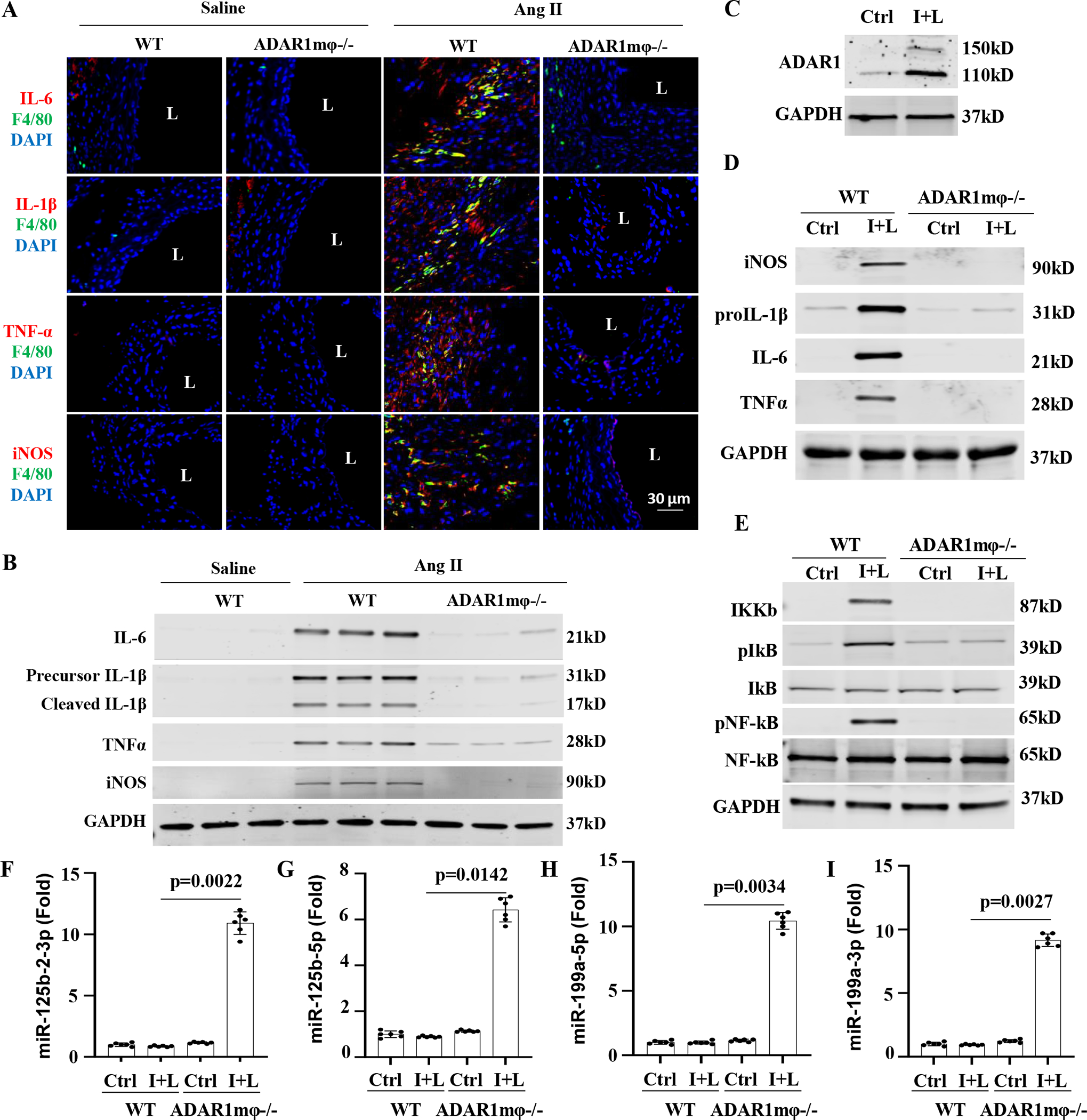
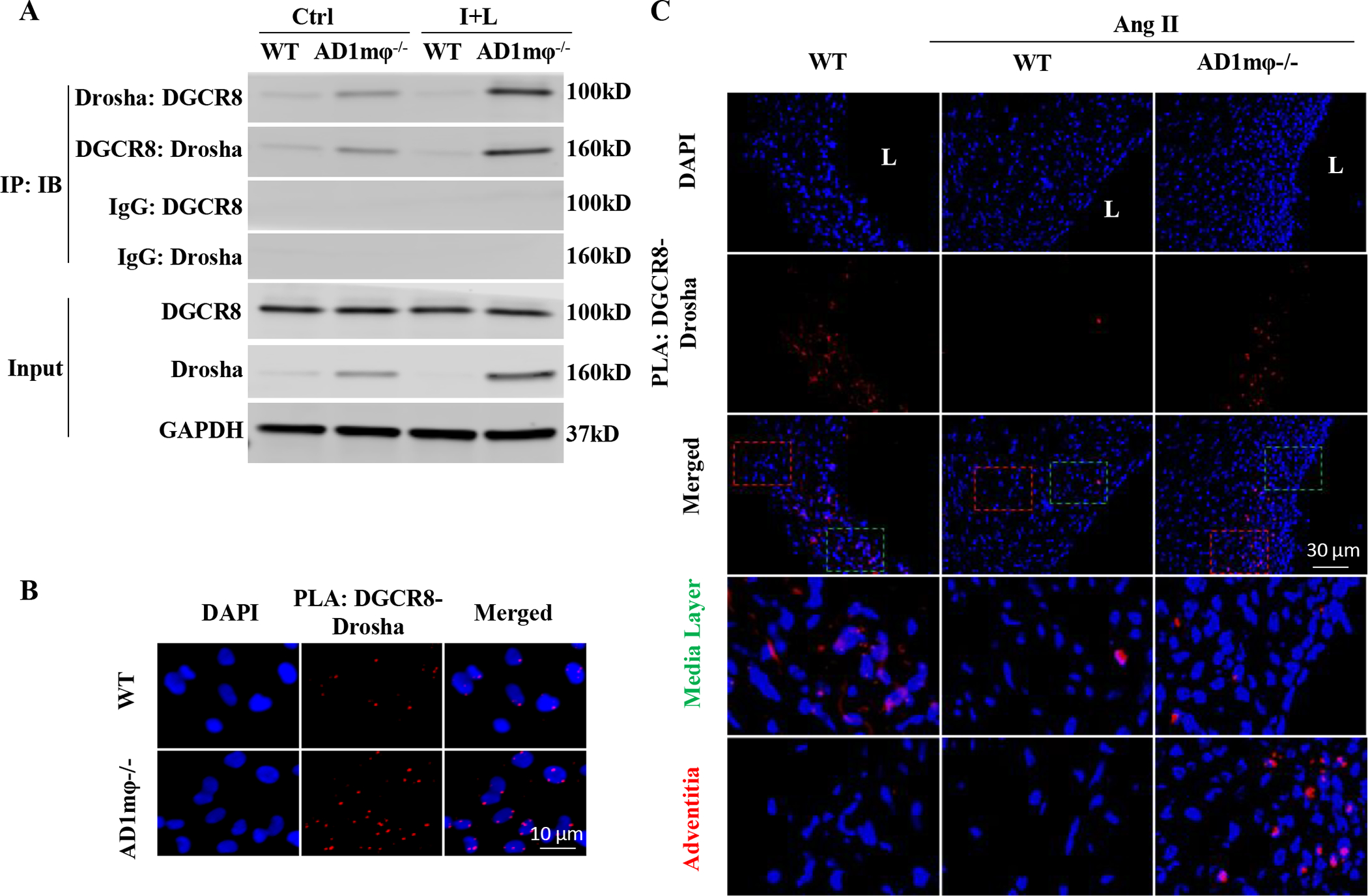
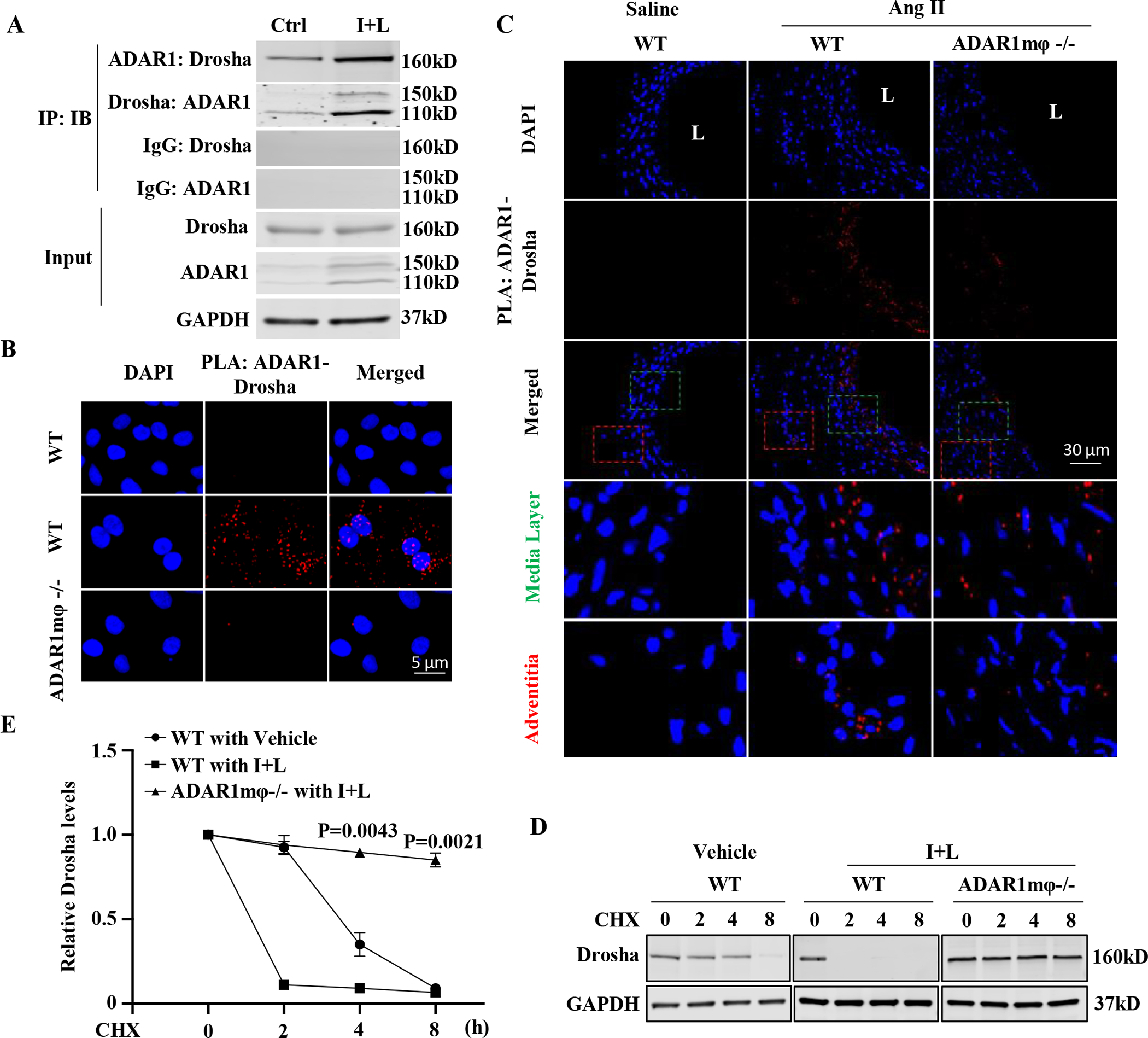
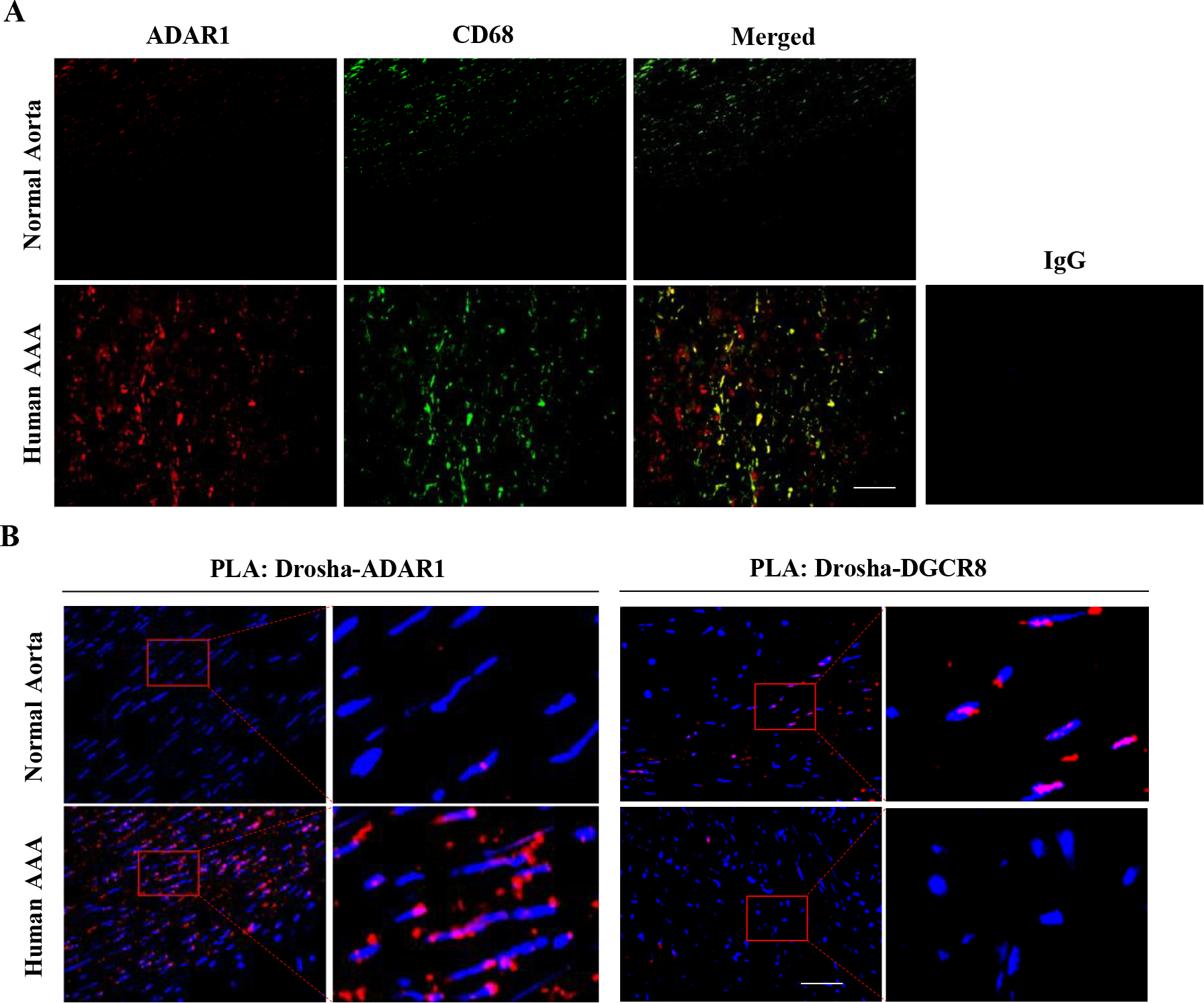
Results: Allograft transplantation of wild-type abdominal aortas to ADAR1+/- recipient mice significantly attenuated AAA formation, suggesting that nonvascular ADAR1 is essential for AAA development. ADAR1 deficiency in hematopoietic cells decreased the prevalence and severity of AAA while inhibited macrophage infiltration and aorta wall inflammation. ADAR1 deletion blocked the classic macrophage activation, diminished NF-κB (nuclear factor kappa B) signaling, and enhanced the expression of a number of anti-inflammatory microRNAs. Mechanistically, ADAR1 interacted with Drosha to promote its degradation, which attenuated Drosha-DGCR8 (DiGeorge syndrome critical region 8) interaction, and consequently inhibited pri- to pre-microRNA processing of microRNAs targeting IKKβ, resulting in an increased IKKβ (inhibitor of nuclear factor kappa-B) expression and enhanced NF-κB signaling. Significantly, ADAR1 was induced in macrophages and interacted with Drosha in human AAA lesions. Reconstitution of ADAR1-deficient, but not the wild type, human monocytes to immunodeficient mice blocked the aneurysm formation in transplanted human arteries.
Conclusions: Macrophage ADAR1 promotes aneurysm formation in both mouse and human arteries through a novel mechanism, that is, Drosha protein degradation, which inhibits the processing of microRNAs targeting NF-kB signaling and thus elicits macrophage-mediated vascular inflammation in AAA.
Keywords: abdominal aortic aneurysm; adenosine deaminases acting on RNA1; humanized AAA model; macrophage; microRNA.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
