

Tetraploidy-linked sensitization to CENP-E inhibition in human cells
- PMID: 36688680
- PMCID: PMC10257419
- DOI: 10.1002/1878-0261.13379
Tetraploidy-linked sensitization to CENP-E inhibition in human cells
Abstract
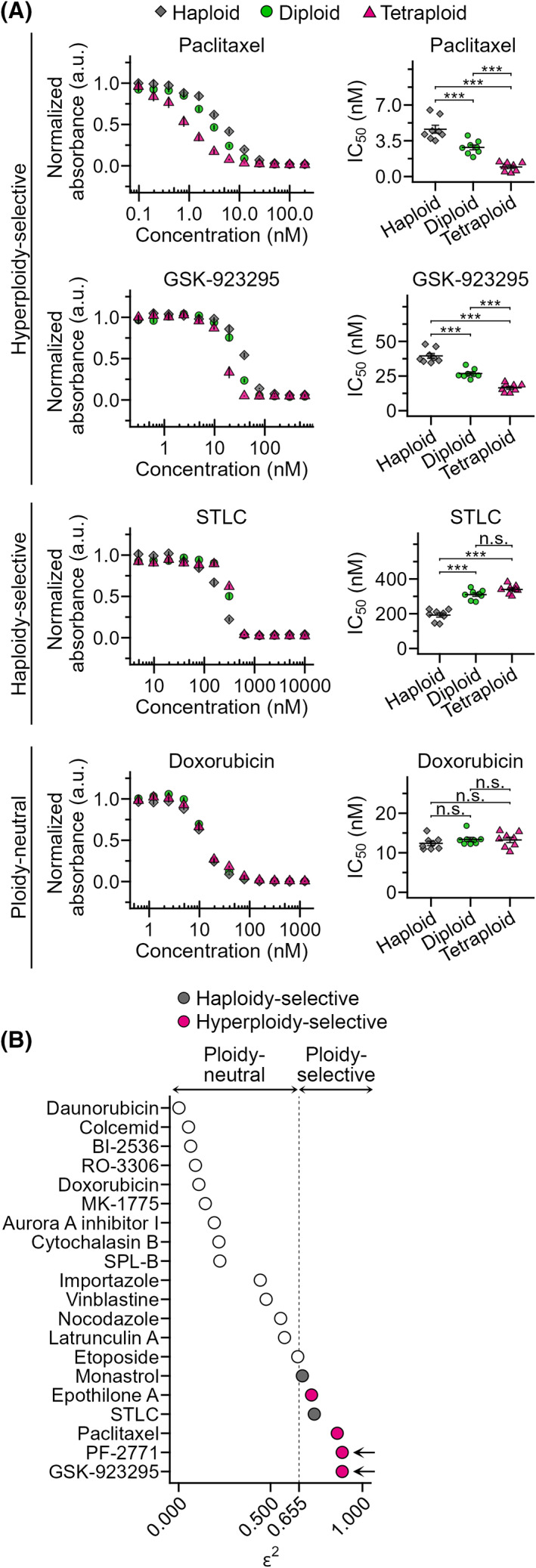
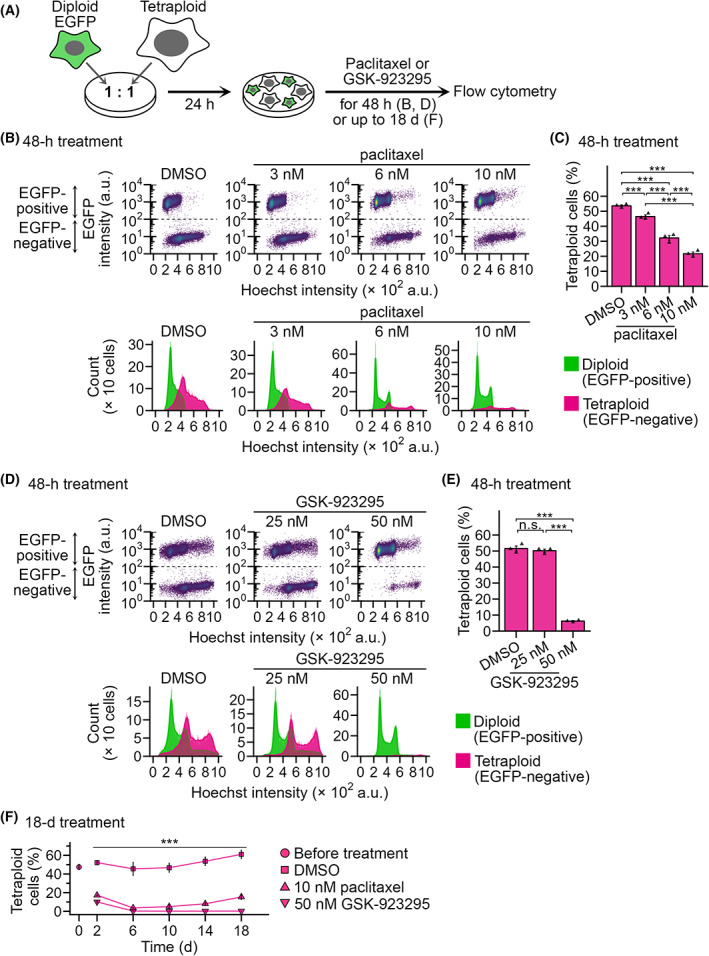
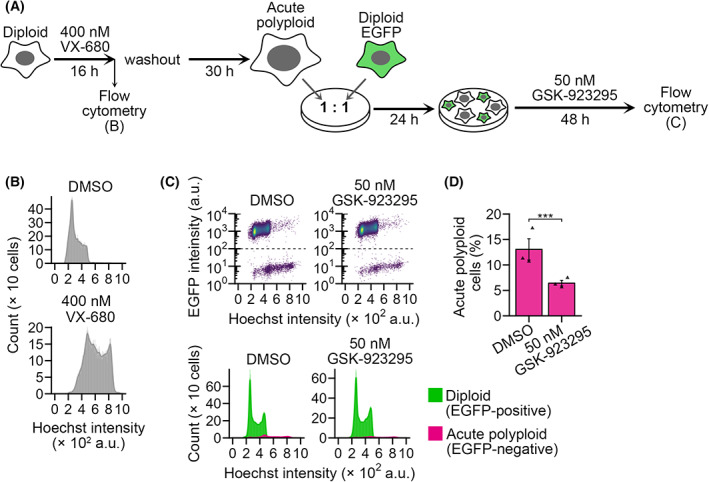
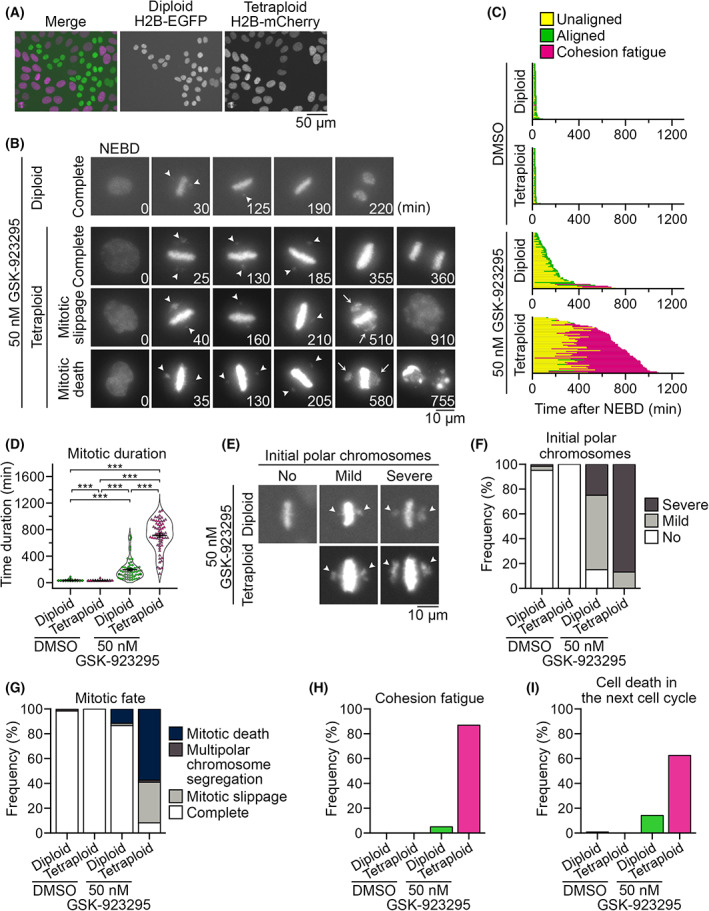
Tetraploidy is a hallmark of cancer cells, and tetraploidy-selective cell growth suppression is a potential strategy for targeted cancer therapy. However, how tetraploid cells differ from normal diploids in their sensitivity to anti-proliferative treatments remains largely unknown. In this study, we found that tetraploid cells are significantly more susceptible to inhibitors of a mitotic kinesin (CENP-E) than are diploids. Treatment with a CENP-E inhibitor preferentially diminished the tetraploid cell population in a diploid-tetraploid co-culture at optimum conditions. Live imaging revealed that a tetraploidy-linked increase in unsolvable chromosome misalignment caused substantially longer mitotic delay in tetraploids than in diploids upon moderate CENP-E inhibition. This time gap of mitotic arrest resulted in cohesion fatigue and subsequent cell death, specifically in tetraploids, leading to tetraploidy-selective cell growth suppression. In contrast, the microtubule-stabilizing compound paclitaxel caused tetraploidy-selective suppression through the aggravation of spindle multipolarization. We also found that treatment with a CENP-E inhibitor had superior generality to paclitaxel in its tetraploidy selectivity across a broader spectrum of cell lines. Our results highlight the unique properties of CENP-E inhibitors in tetraploidy-selective suppression and their potential use in the development of tetraploidy-targeting interventions in cancer.
Keywords: chromosome; mitosis; motor protein; ploidy.
© 2023 The Authors. Molecular Oncology published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Preferential Killing of Tetraploid Colon Cancer Cells by Targeting the Mitotic Kinase PLK1.Cell Physiol Biochem. 2020 Apr 8;54(2):303-320. doi: 10.33594/000000221. Cell Physiol Biochem. 2020. PMID: 32259417
-
CENP-E Inhibition Induces Chromosomal Instability and Synergizes with Diverse Microtubule-Targeting Agents in Breast Cancer.Cancer Res. 2024 Aug 15;84(16):2674-2689. doi: 10.1158/0008-5472.CAN-23-3332. Cancer Res. 2024. PMID: 38832939 Free PMC article.
-
Mitosis. Microtubule detyrosination guides chromosomes during mitosis.Science. 2015 May 15;348(6236):799-803. doi: 10.1126/science.aaa5175. Epub 2015 Apr 23. Science. 2015. PMID: 25908662 Free PMC article.
-
Leaving no-one behind: how CENP-E facilitates chromosome alignment.Essays Biochem. 2020 Sep 4;64(2):313-324. doi: 10.1042/EBC20190073. Essays Biochem. 2020. PMID: 32347304 Free PMC article. Review.
-
CENP-E as a target for cancer therapy: Where are we now?Life Sci. 2018 Sep 1;208:192-200. doi: 10.1016/j.lfs.2018.07.037. Epub 2018 Jul 19. Life Sci. 2018. PMID: 30031812 Review.
Cited by
-
Kinesin-7 CENP-E in tumorigenesis: Chromosome instability, spindle assembly checkpoint, and applications.Front Mol Biosci. 2024 Mar 15;11:1366113. doi: 10.3389/fmolb.2024.1366113. eCollection 2024. Front Mol Biosci. 2024. PMID: 38560520 Free PMC article. Review.
-
Comparative Pharmacological Analysis of Mitotic Inhibitors Using Isogenic Ploidy Series of HAP1 Cells.Methods Mol Biol. 2025;2872:207-219. doi: 10.1007/978-1-0716-4224-5_14. Methods Mol Biol. 2025. PMID: 39616578
-
Photochemical Control of Cell Division Using a Photoswitchable CENP-E Inhibitor.Methods Mol Biol. 2025;2872:101-113. doi: 10.1007/978-1-0716-4224-5_7. Methods Mol Biol. 2025. PMID: 39616571
-
Tetraploidy in normal tissues and diseases: mechanisms and consequences.Chromosoma. 2025 Mar 21;134(1):3. doi: 10.1007/s00412-025-00829-1. Chromosoma. 2025. PMID: 40117022 Free PMC article. Review.
-
Targeting Kinesins for Therapeutic Exploitation of Chromosomal Instability in Lung Cancer.Cancers (Basel). 2025 Feb 18;17(4):685. doi: 10.3390/cancers17040685. Cancers (Basel). 2025. PMID: 40002279 Free PMC article. Review.
References
-
- Fujiwara T, Bandi M, Nitta M, Ivanova EV, Bronson RT, Pellman D. Cytokinesis failure generating tetraploids promotes tumorigenesis in p53‐null cells. Nature. 2005;437:1043–7. - PubMed
MeSH terms
Substances
Grants and funding
- Daiichi Sankyo Foundation of Life Science
- Hokkaido University-Hitachi Joint Cooperative Support Program for Education and Research
- JP19H03219/Japan Society for the Promotion of Science
- JP19H05413/Japan Society for the Promotion of Science
- JP19J12210/Japan Society for the Promotion of Science
- JP19KK0181/Japan Society for the Promotion of Science
- JP21K19244/Japan Society for the Promotion of Science
- JP21K20737/Japan Society for the Promotion of Science
- JPJSBP120193801/Japan Society for the Promotion of Science
- The Kato Memorial Bioscience Foundation
- The Nakatani Foundation
- The Orange Foundation
- The Princess Takamatsu Cancer Research Fund
- the Sasakawa Scientific Research Grant
- The Smoking Research Foundation
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
