

Biogenesis of JC polyomavirus associated extracellular vesicles
- PMID: 36688929
- PMCID: PMC9854252
- DOI: 10.1002/jex2.43
Biogenesis of JC polyomavirus associated extracellular vesicles
Abstract
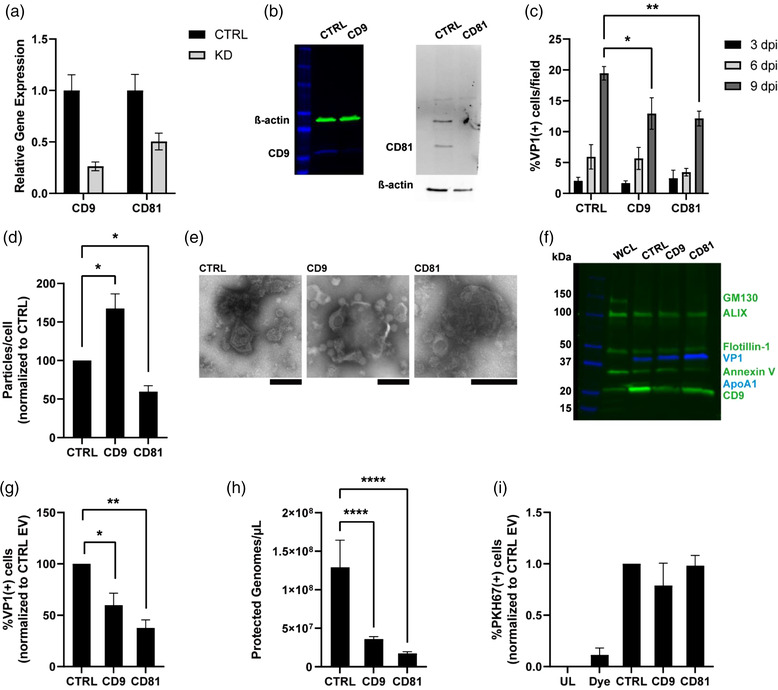
JC polyomavirus (JCPyV) is a small, non-enveloped virus that persists in the kidney in about half the adult population. In severely immune-compromised individuals JCPyV causes the neurodegenerative disease progressive multifocal leukoencephalopathy (PML) in the brain. JCPyV has been shown to infect cells by both direct and indirect mechanisms, the latter involving extracellular vesicle (EV) mediated infection. While direct mechanisms of infection are well studied indirect EV mediated mechanisms are poorly understood. Using a combination of chemical and genetic approaches we show that several overlapping intracellular pathways are responsible for the biogenesis of virus containing EV. Here we show that targeting neutral sphingomyelinase 2 (nSMase2) with the drug cambinol decreased the spread of JCPyV over several viral life cycles. Genetic depletion of nSMase2 by either shRNA or CRISPR/Cas9 reduced EV-mediated infection. Individual knockdown of seven ESCRT-related proteins including HGS, ALIX, TSG101, VPS25, VPS20, CHMP4A, and VPS4A did not significantly reduce JCPyV associated EV (JCPyV(+) EV) infectivity, whereas knockdown of the tetraspanins CD9 and CD81 or trafficking and/or secretory autophagy-related proteins RAB8A, RAB27A, and GRASP65 all significantly reduced the spread of JCPyV and decreased EV-mediated infection. These findings point to a role for exosomes and secretory autophagosomes in the biogenesis of JCPyV associated EVs with specific roles for nSMase2, CD9, CD81, RAB8A, RAB27A, and GRASP65 proteins.
Keywords: JC polyomavirus; biogenesis; extracellular vesicles.
Conflict of interest statement
CONFLICT OF INTEREST DISCLOSURE No conflicts of interest are declared by the authors.
Figures




References
-
- Alemu, E. A. , Lamark, T. , Torgersen, K. M. , Birgisdottir, A. B. , Larsen, K. B. , Jain, A. , Olsvik, H. , Øvervatn, A. , Kirkin, V. , & Johansen, T. (2012). ATG8 family proteins act as scaffolds for assembly of the ULK complex: Sequence requirements for LC3‐interacting region (LIR) motifs. Journal of Biological Chemistry, 287(47), 39275–39290. - PMC - PubMed
-
- Altan‐Bonnet, N. , Perales, C. , & Domingo, E. (2019). Extracellular vesicles: Vehicles of en bloc viral transmission. Virus Research, 265, 143–149. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous