

Yeast derlin Dfm1 employs a chaperone-like function to resolve misfolded membrane protein stress
- PMID: 36689475
- PMCID: PMC9894555
- DOI: 10.1371/journal.pbio.3001950
Yeast derlin Dfm1 employs a chaperone-like function to resolve misfolded membrane protein stress
Abstract
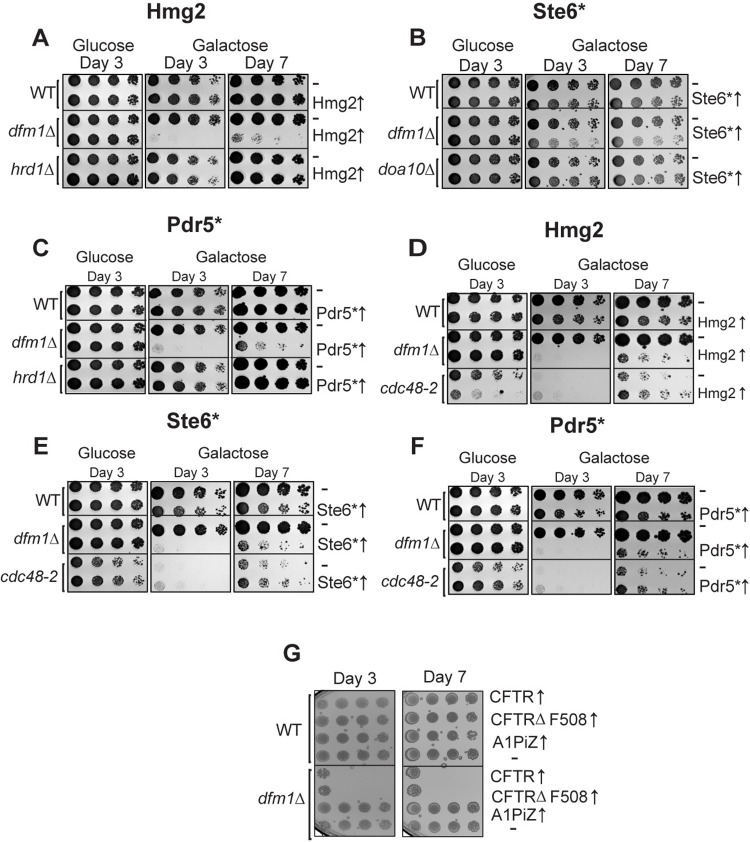
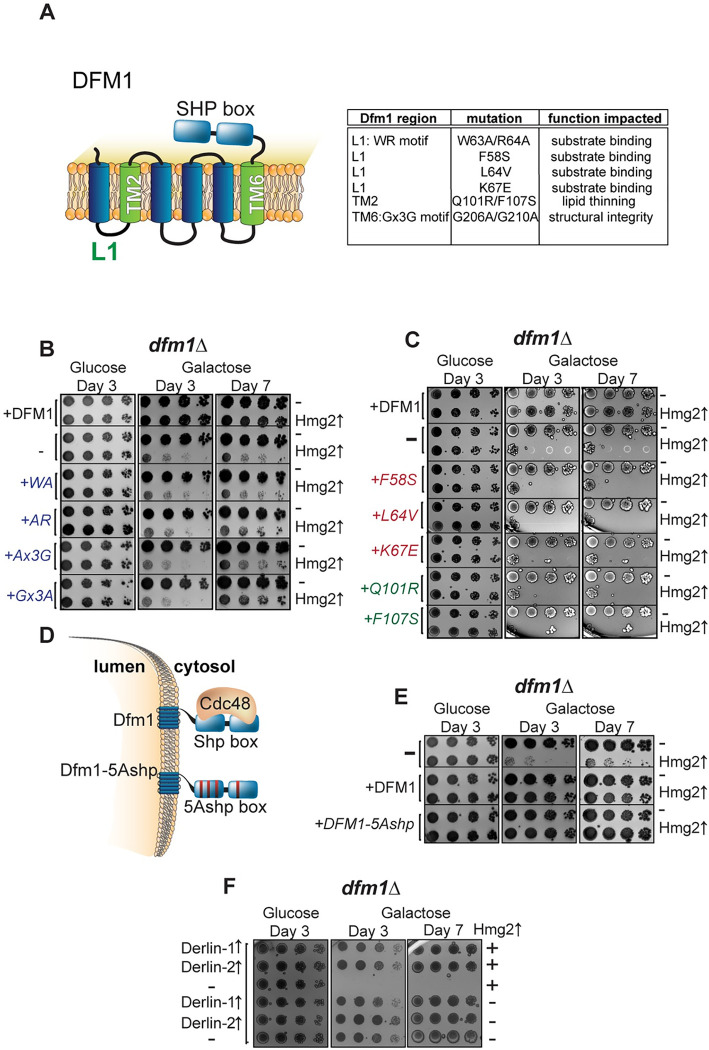
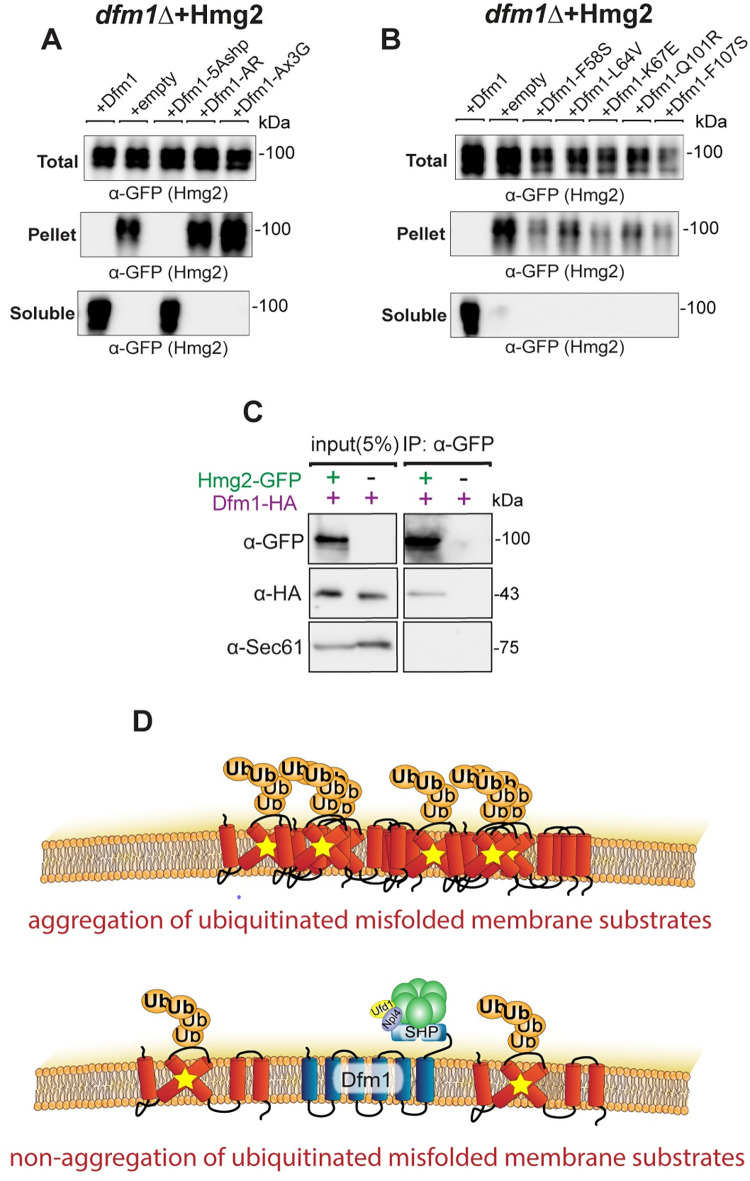
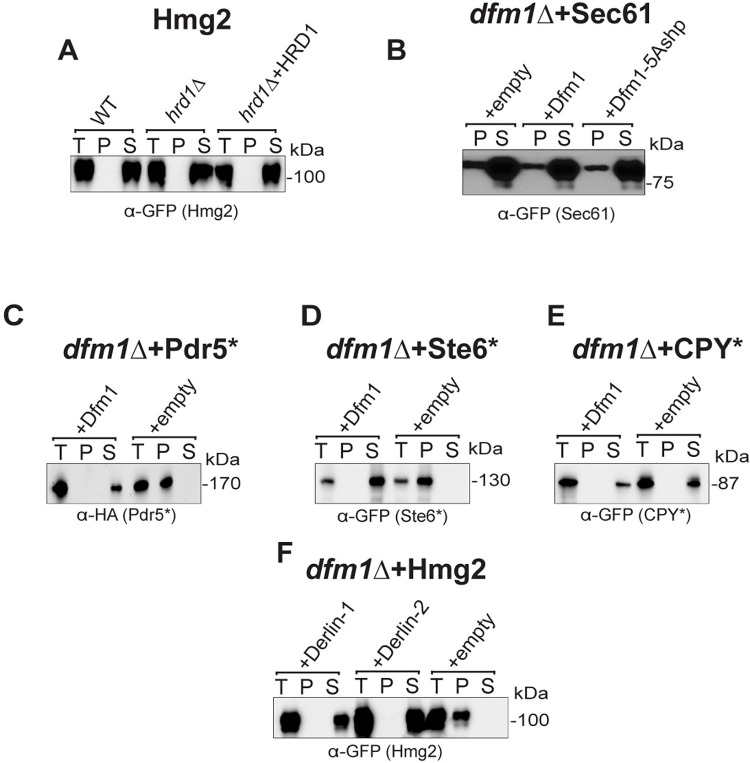
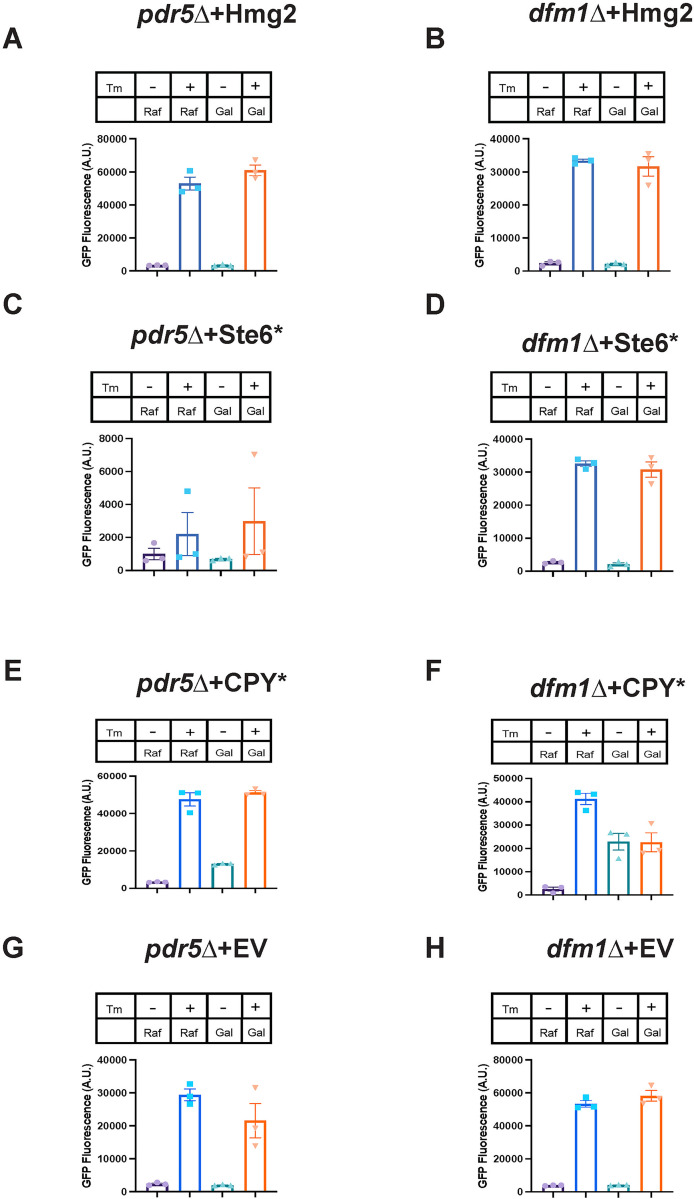
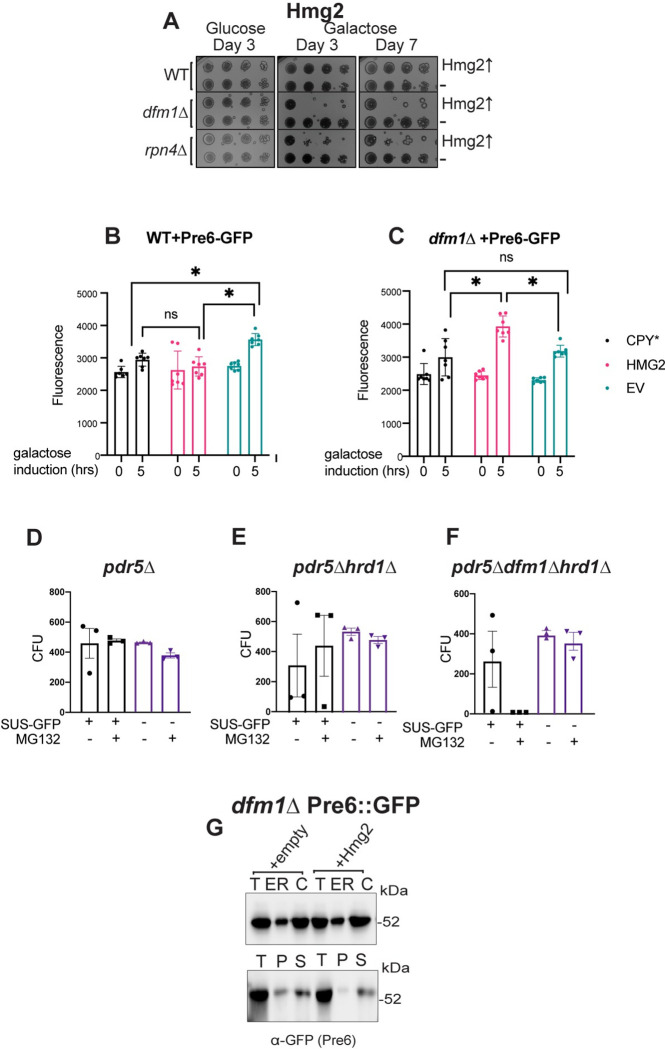
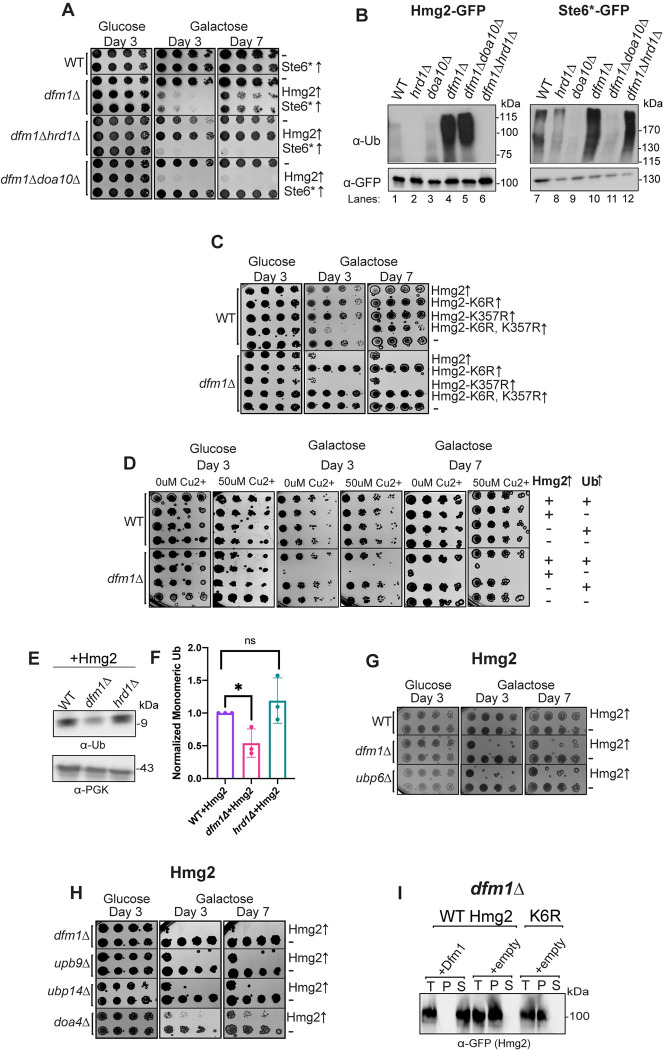
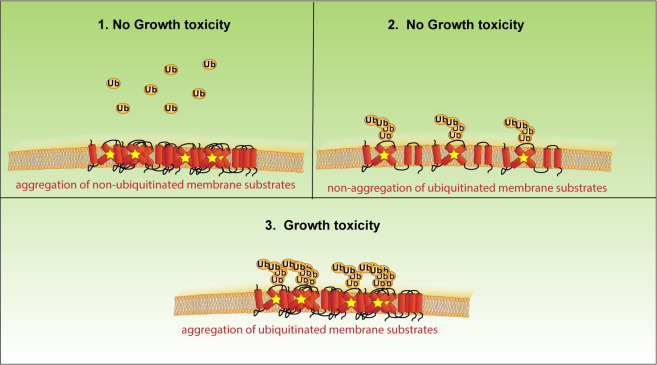
Protein aggregates are a common feature of diseased and aged cells. Membrane proteins comprise a quarter of the proteome, and yet, it is not well understood how aggregation of membrane proteins is regulated and what effects these aggregates can have on cellular health. We have determined in yeast that the derlin Dfm1 has a chaperone-like activity that influences misfolded membrane protein aggregation. We establish that this function of Dfm1 does not require recruitment of the ATPase Cdc48 and it is distinct from Dfm1's previously identified function in dislocating misfolded membrane proteins from the endoplasmic reticulum (ER) to the cytosol for degradation. Additionally, we assess the cellular impacts of misfolded membrane proteins in the absence of Dfm1 and determine that misfolded membrane proteins are toxic to cells in the absence of Dfm1 and cause disruptions to proteasomal and ubiquitin homeostasis.
Copyright: © 2023 Kandel et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared no competing interests exist.
Figures
Similar articles
-
The derlin Dfm1 couples retrotranslocation of a folded protein domain to its proteasomal degradation.J Cell Biol. 2024 May 6;223(5):e202308074. doi: 10.1083/jcb.202308074. Epub 2024 Mar 5. J Cell Biol. 2024. PMID: 38448163 Free PMC article.
-
The Dfm1 Derlin Is Required for ERAD Retrotranslocation of Integral Membrane Proteins.Mol Cell. 2018 Jan 18;69(2):306-320.e4. doi: 10.1016/j.molcel.2017.12.012. Mol Cell. 2018. PMID: 29351849 Free PMC article.
-
Derlin rhomboid pseudoproteases employ substrate engagement and lipid distortion to enable the retrotranslocation of ERAD membrane substrates.Cell Rep. 2021 Oct 19;37(3):109840. doi: 10.1016/j.celrep.2021.109840. Cell Rep. 2021. PMID: 34686332 Free PMC article.
-
Protein Quality Control of the Endoplasmic Reticulum and Ubiquitin-Proteasome-Triggered Degradation of Aberrant Proteins: Yeast Pioneers the Path.Annu Rev Biochem. 2018 Jun 20;87:751-782. doi: 10.1146/annurev-biochem-062917-012749. Epub 2018 Feb 2. Annu Rev Biochem. 2018. PMID: 29394096 Review.
-
Endoplasmic reticulum associated protein degradation: a chaperone assisted journey to hell.Biochim Biophys Acta. 2010 Jun;1803(6):694-705. doi: 10.1016/j.bbamcr.2010.02.005. Epub 2010 Feb 25. Biochim Biophys Acta. 2010. PMID: 20219571 Review.
Cited by
-
Multiomics of GCN4-Dependent Replicative Lifespan Extension Models Reveals Gcn4 as a Regulator of Protein Turnover in Yeast.Int J Mol Sci. 2023 Nov 10;24(22):16163. doi: 10.3390/ijms242216163. Int J Mol Sci. 2023. PMID: 38003352 Free PMC article.
-
The derlin Dfm1 couples retrotranslocation of a folded protein domain to its proteasomal degradation.J Cell Biol. 2024 May 6;223(5):e202308074. doi: 10.1083/jcb.202308074. Epub 2024 Mar 5. J Cell Biol. 2024. PMID: 38448163 Free PMC article.
-
Selenoprotein K at the intersection of cellular pathways.Arch Biochem Biophys. 2025 Feb;764:110221. doi: 10.1016/j.abb.2024.110221. Epub 2024 Nov 20. Arch Biochem Biophys. 2025. PMID: 39571956 Review.
-
Proteotoxic stress and the ubiquitin proteasome system.Semin Cell Dev Biol. 2024 Mar 15;156:107-120. doi: 10.1016/j.semcdb.2023.08.002. Epub 2023 Sep 19. Semin Cell Dev Biol. 2024. PMID: 37734998 Free PMC article. Review.
-
An ERAD-independent role for rhomboid pseudoprotease Dfm1 in mediating sphingolipid homeostasis.EMBO J. 2023 Feb 15;42(4):e112275. doi: 10.15252/embj.2022112275. Epub 2022 Nov 9. EMBO J. 2023. PMID: 36350249 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
