

Touch neurons underlying dopaminergic pleasurable touch and sexual receptivity
- PMID: 36693373
- PMCID: PMC9898224
- DOI: 10.1016/j.cell.2022.12.034
Touch neurons underlying dopaminergic pleasurable touch and sexual receptivity
Abstract
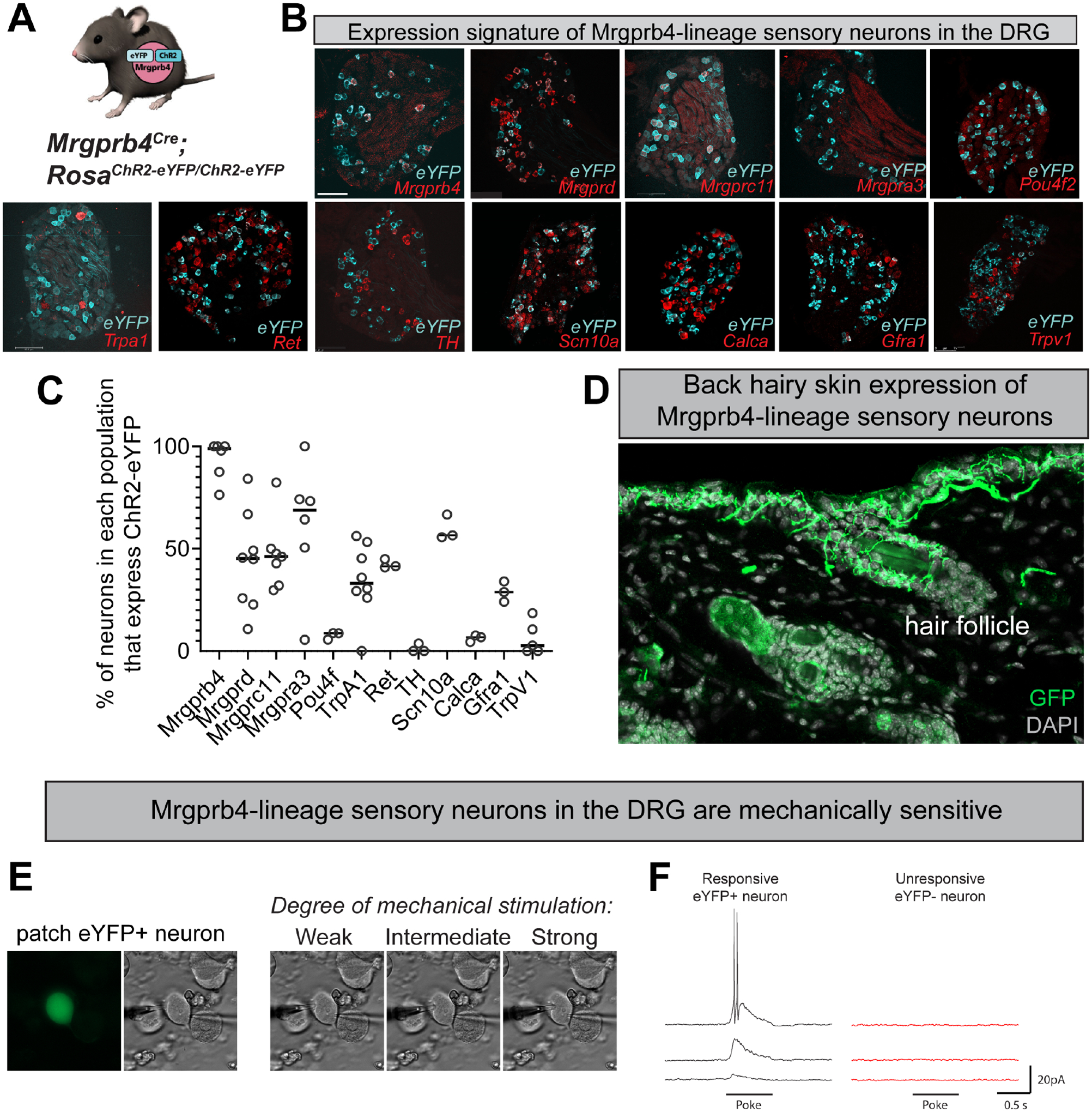
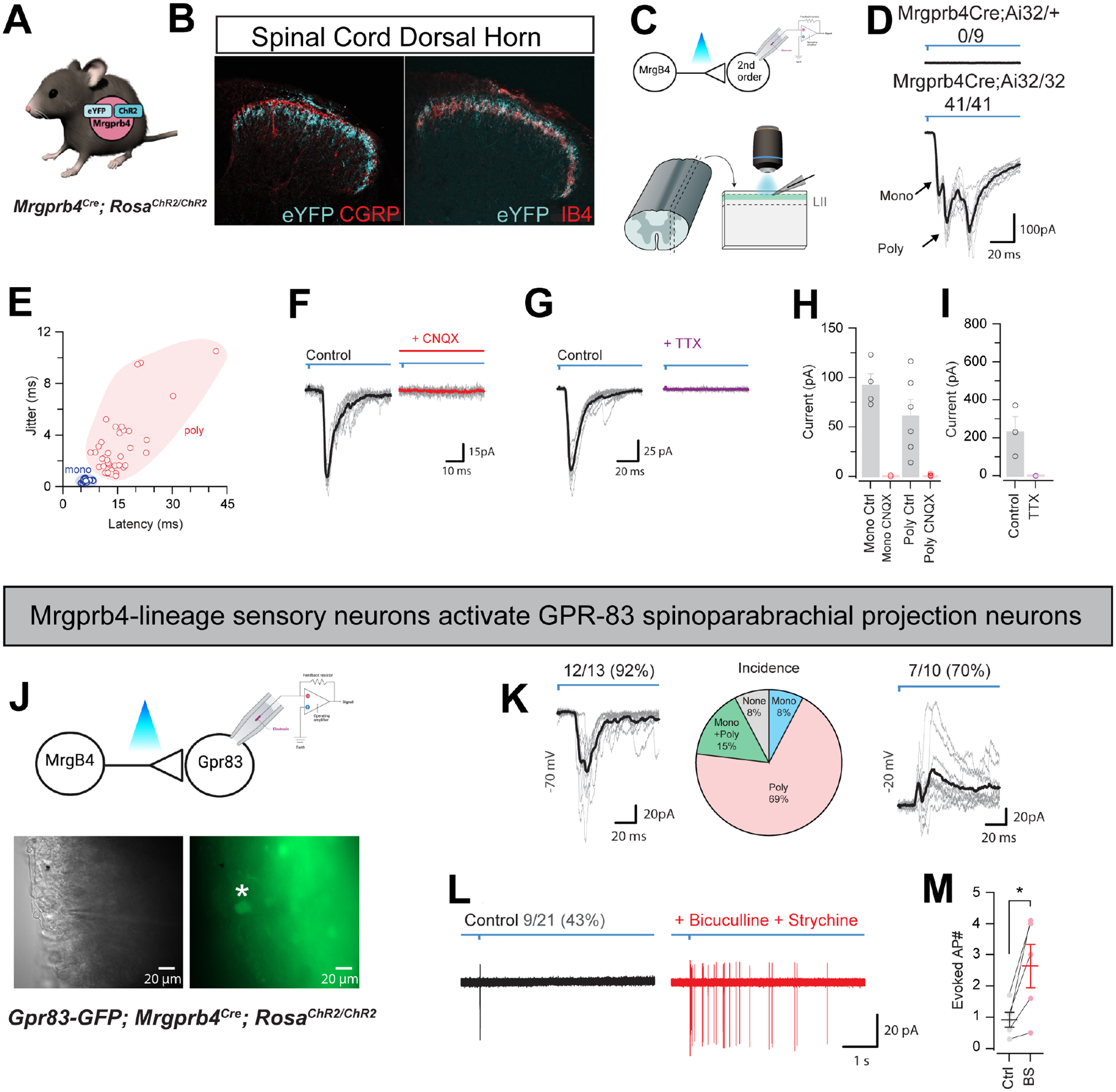
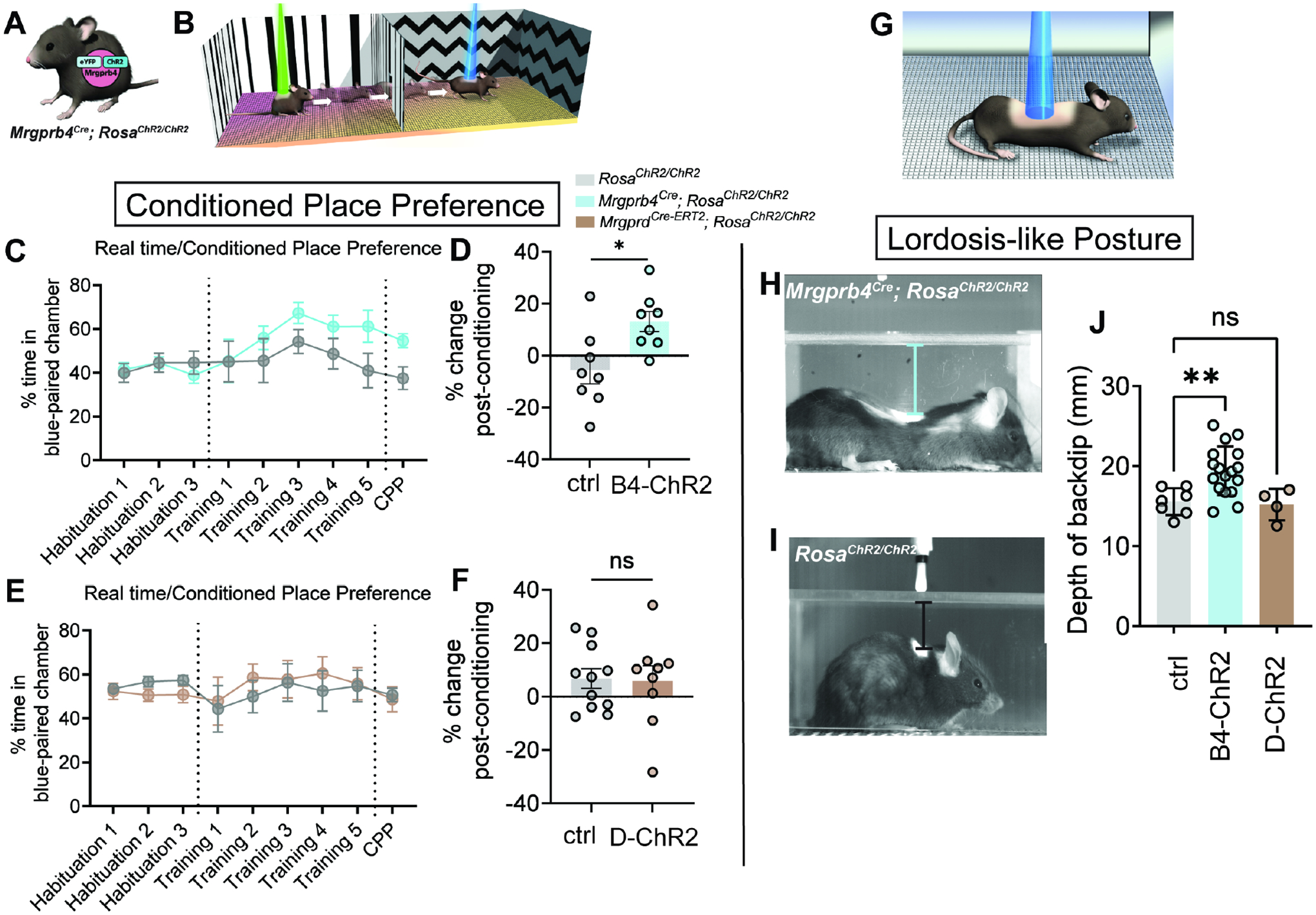
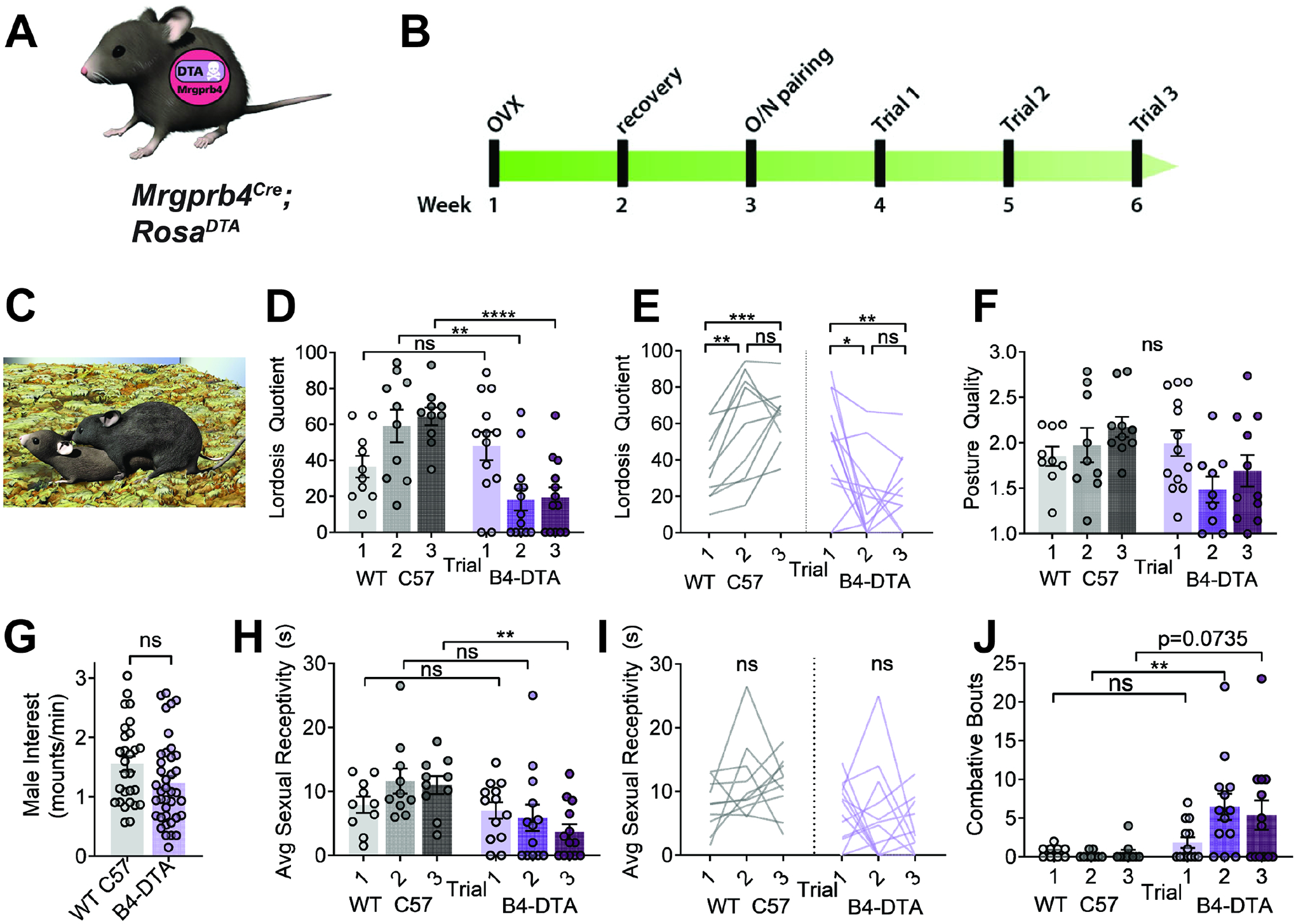
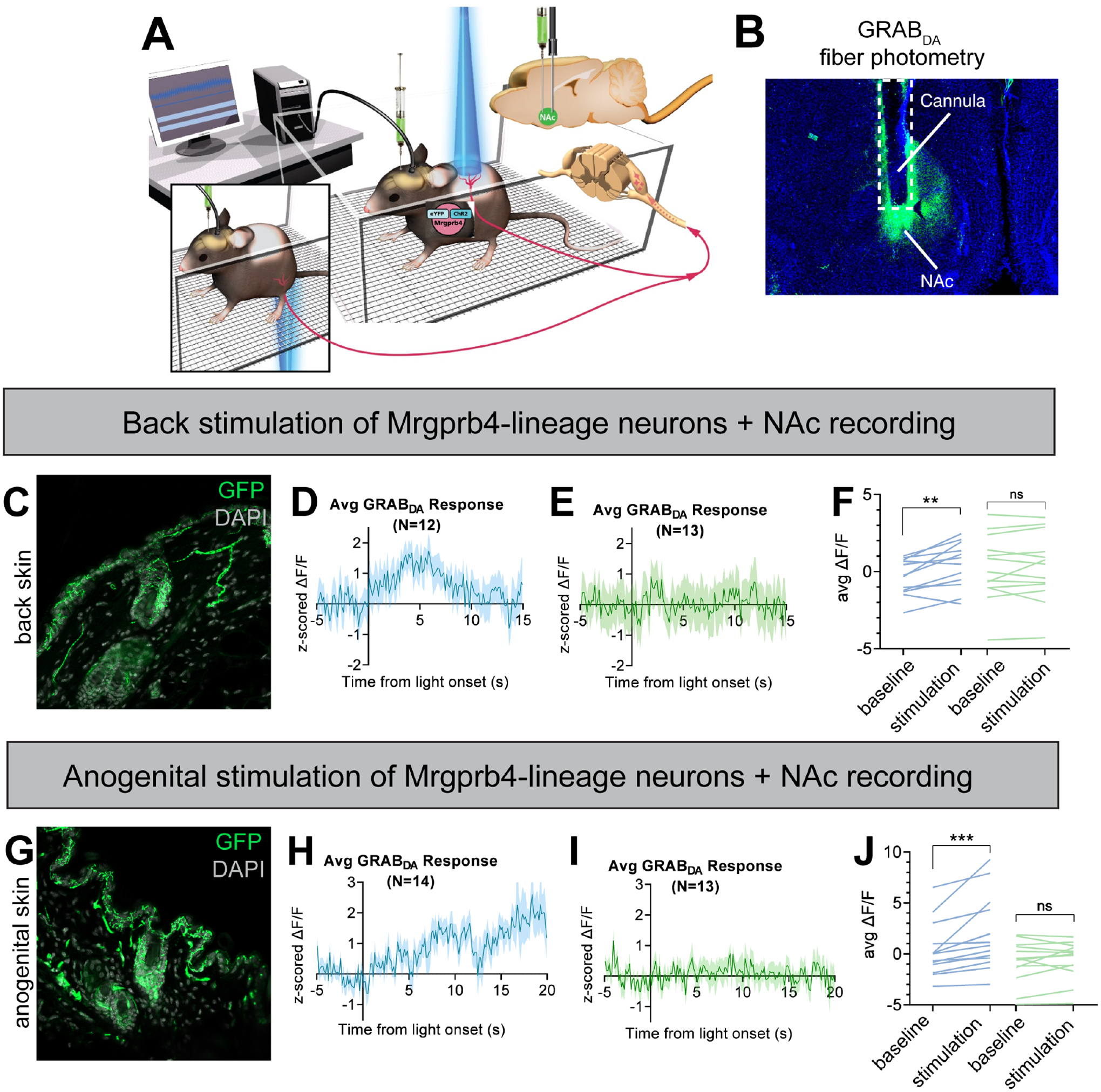
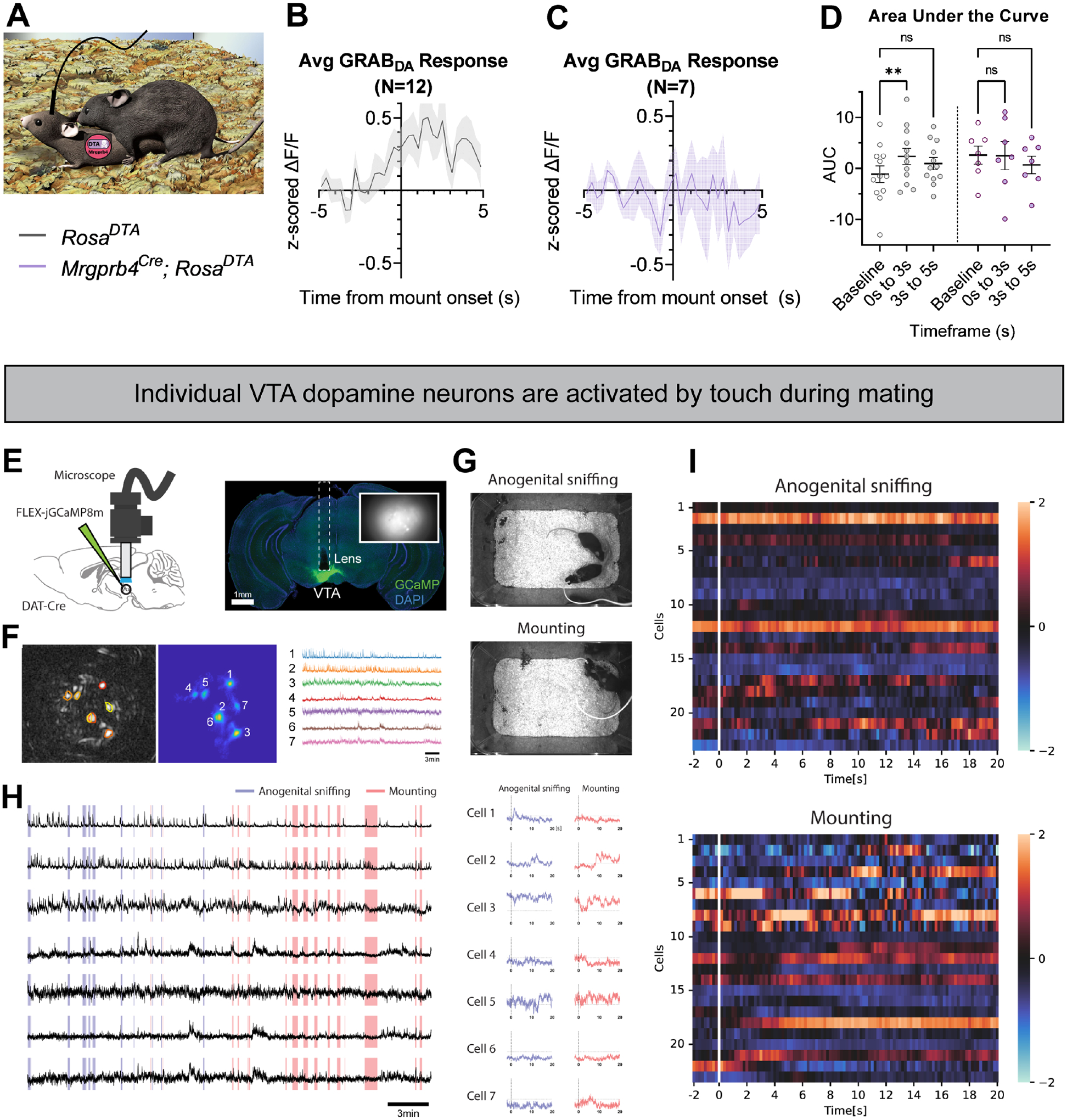
Pleasurable touch is paramount during social behavior, including sexual encounters. However, the identity and precise role of sensory neurons that transduce sexual touch remain unknown. A population of sensory neurons labeled by developmental expression of the G protein-coupled receptor Mrgprb4 detects mechanical stimulation in mice. Here, we study the social relevance of Mrgprb4-lineage neurons and reveal that these neurons are required for sexual receptivity and sufficient to induce dopamine release in the brain. Even in social isolation, optogenetic stimulation of Mrgprb4-lineage neurons through the back skin is sufficient to induce a conditioned place preference and a striking dorsiflexion resembling the lordotic copulatory posture. In the absence of Mrgprb4-lineage neurons, female mice no longer find male mounts rewarding: sexual receptivity is supplanted by aggression and a coincident decline in dopamine release in the nucleus accumbens. Together, these findings establish that Mrgprb4-lineage neurons initiate a skin-to-brain circuit encoding the rewarding quality of social touch.
Keywords: peripheral optogenetics; pleasure; reward pathway; spinal cord; touch.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Orefice LL, Mosko JR, Morency DT, Wells MF, Tasnim A, Mozeika SM, Ye M, Chirila AM, Emanuel AJ, Rankin G, Fame RM, Lehtinen MK, Feng G, Ginty DD. Targeting Peripheral Somatosensory Neurons to Improve Tactile-Related Phenotypes in ASD Models. Cell. 2019;178(4):867–86 e24. Epub 2019/08/10. doi: 10.1016/j.cell.2019.07.024. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
