

Higher-order SPOP assembly reveals a basis for cancer mutant dysregulation
- PMID: 36693379
- PMCID: PMC9992347
- DOI: 10.1016/j.molcel.2022.12.033
Higher-order SPOP assembly reveals a basis for cancer mutant dysregulation
Abstract
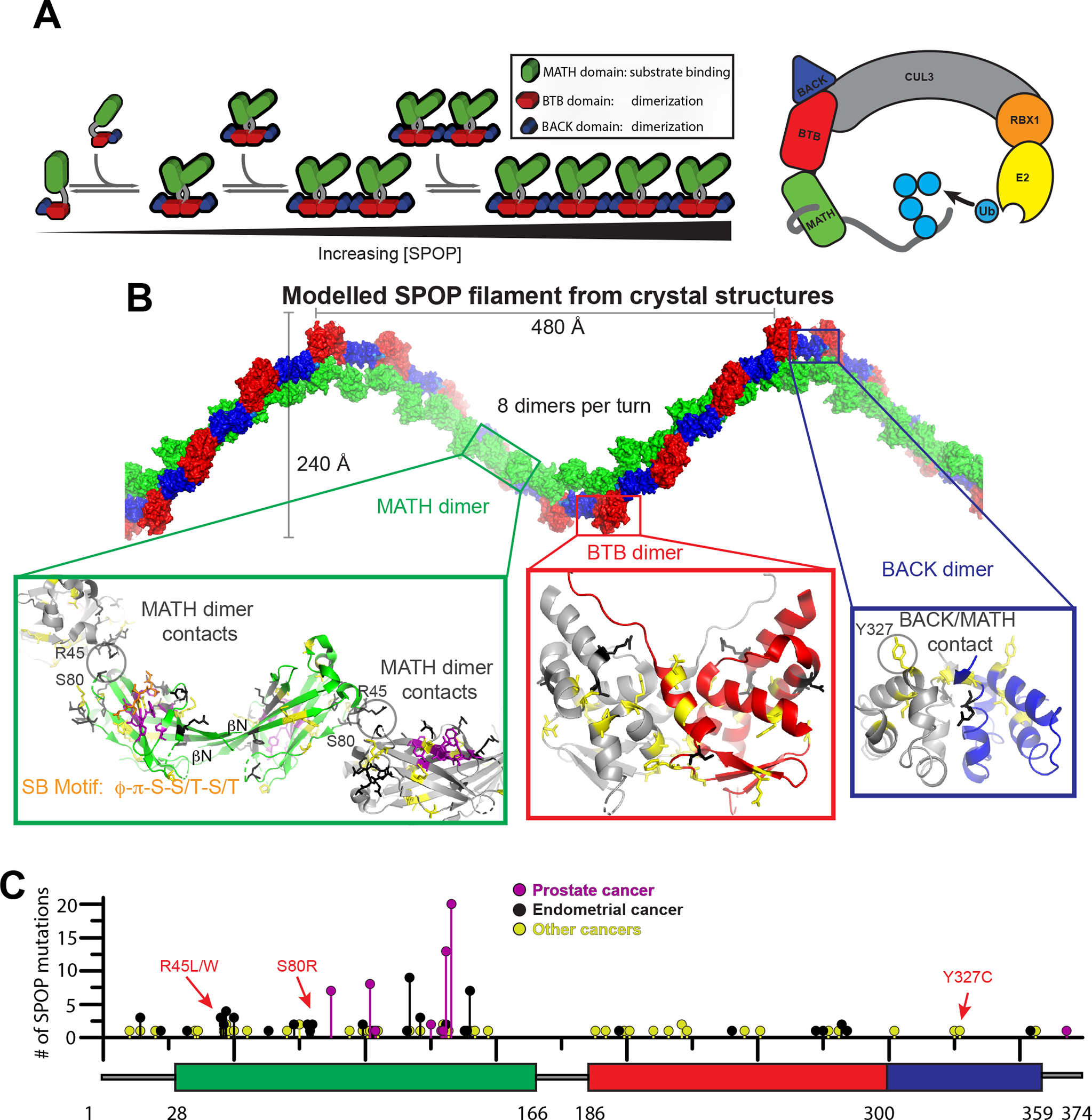
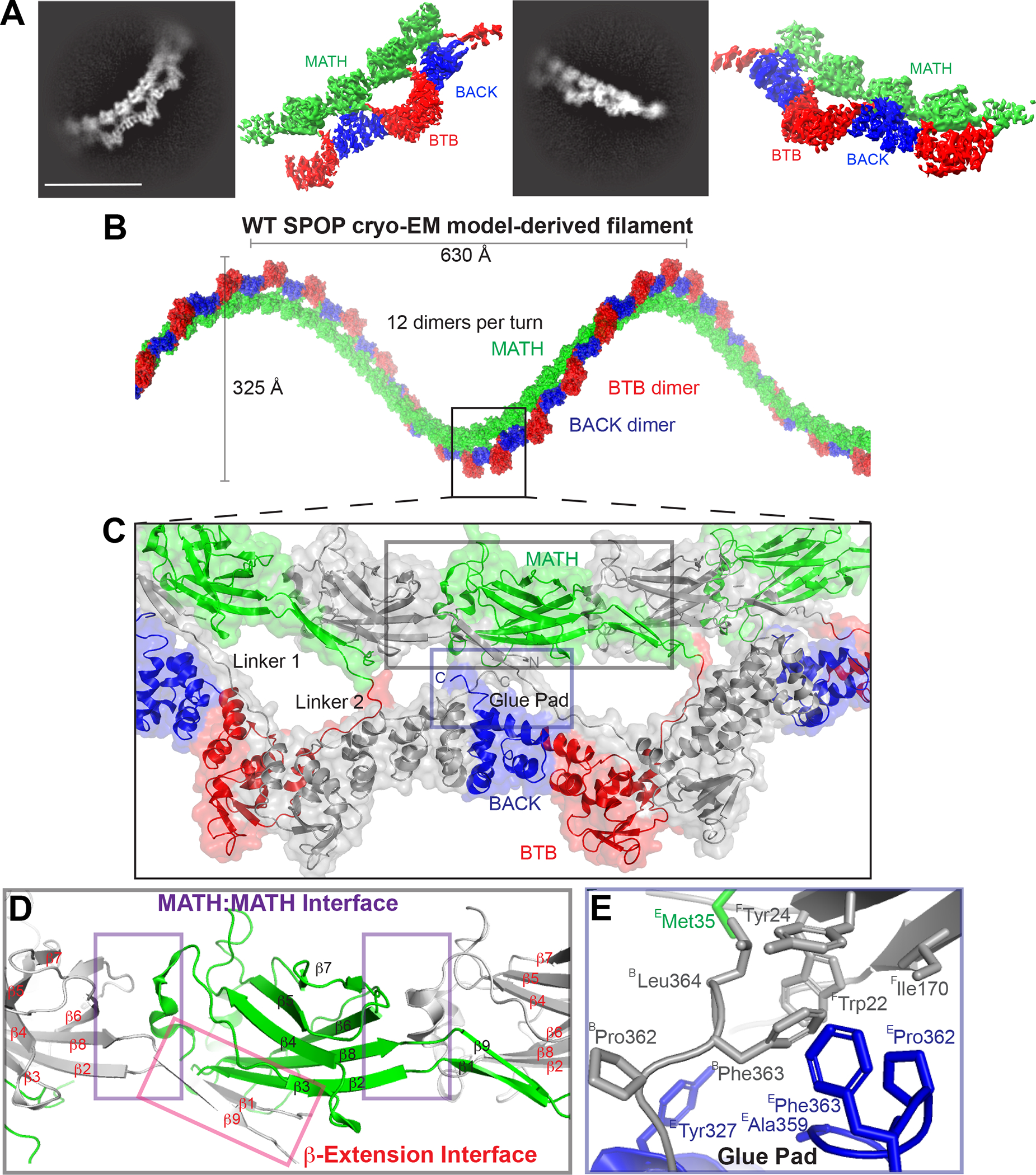
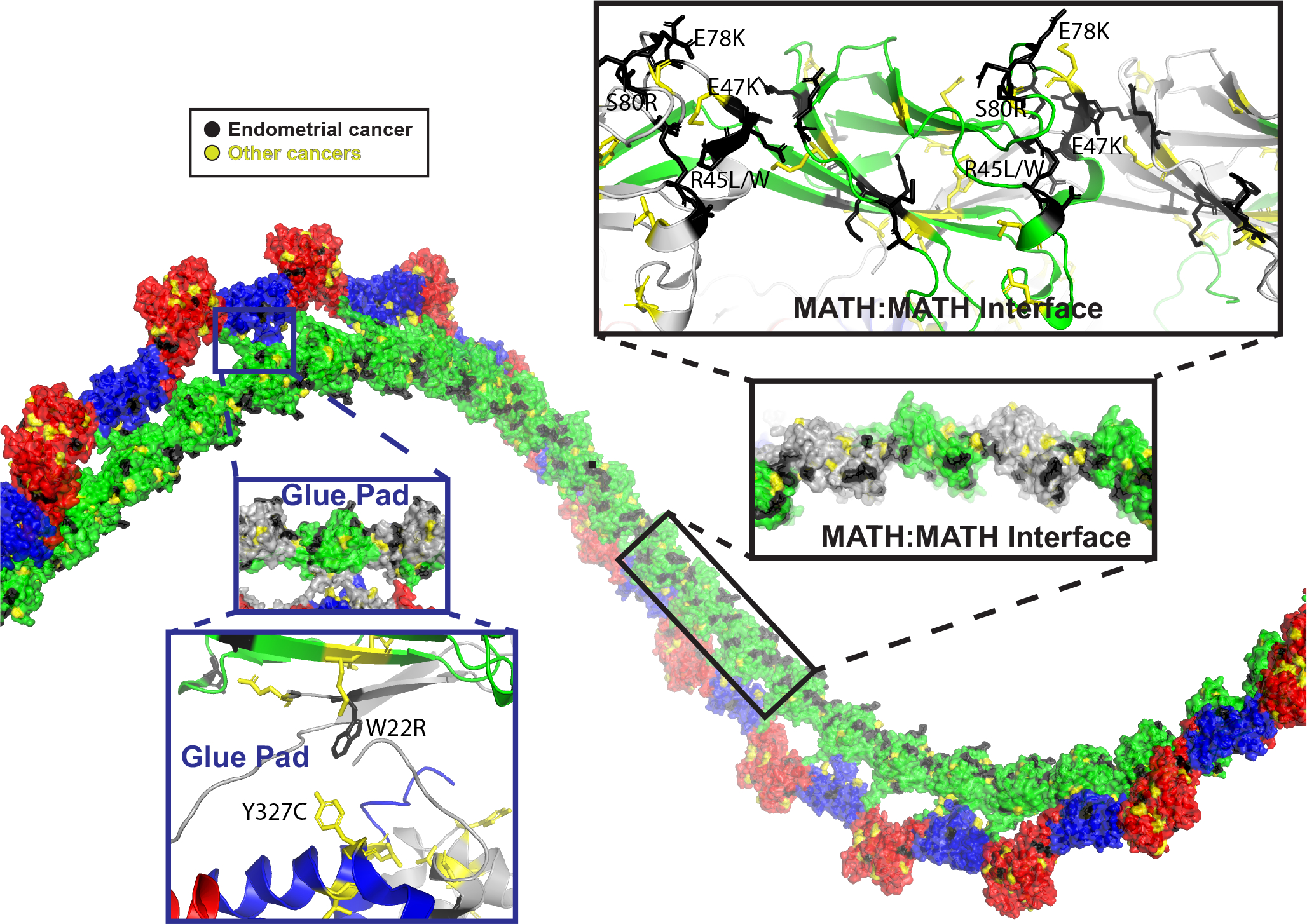
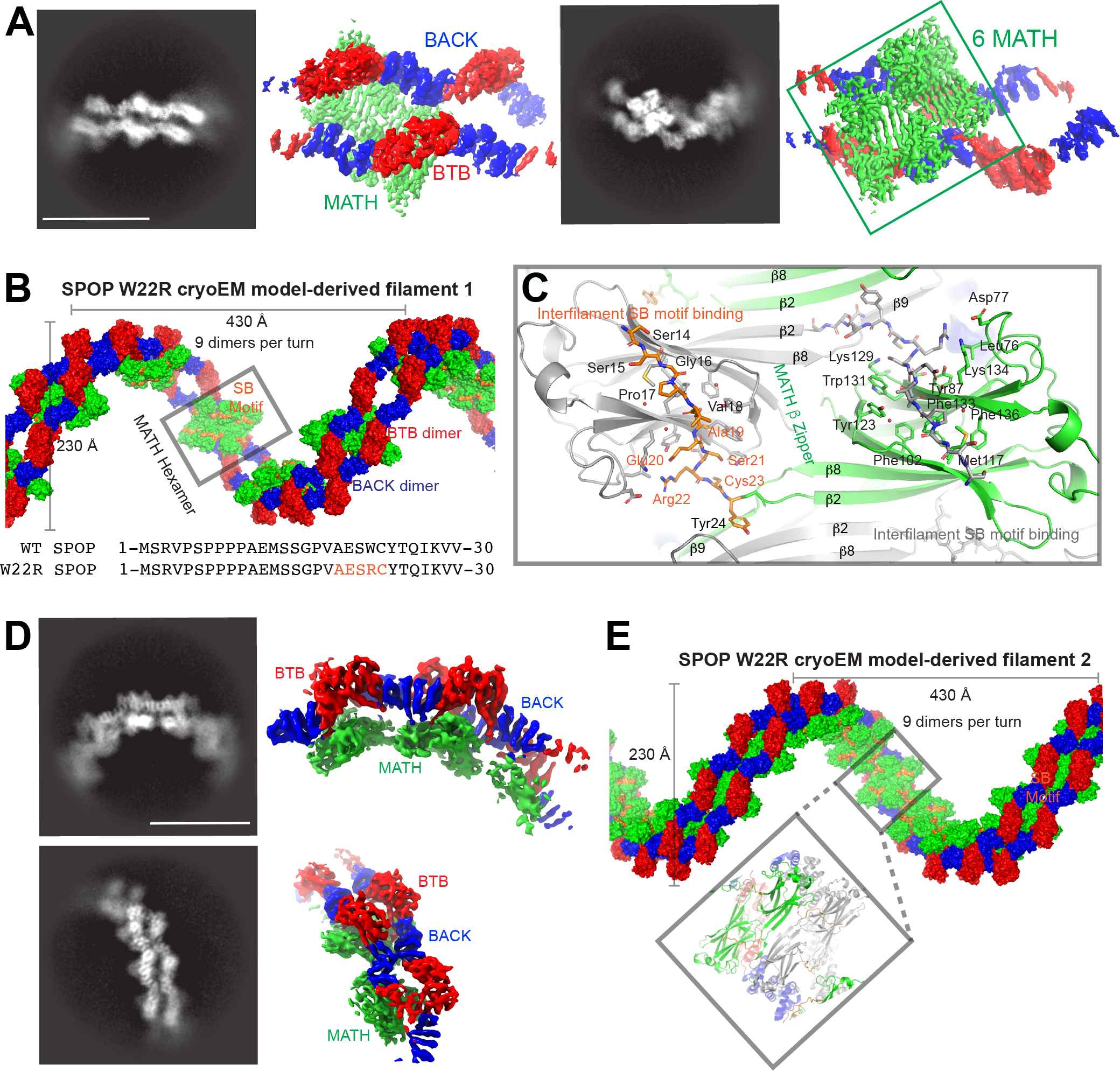
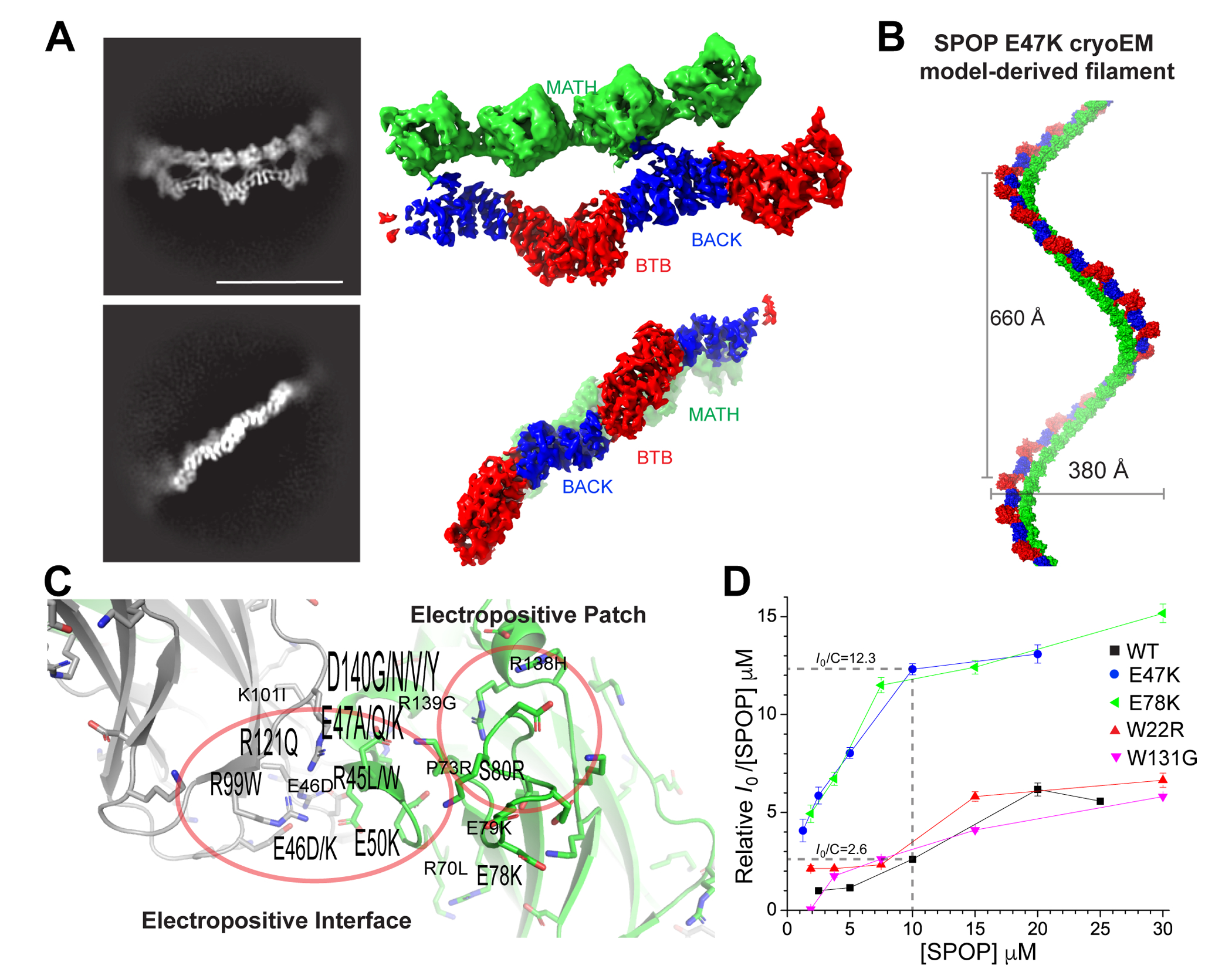
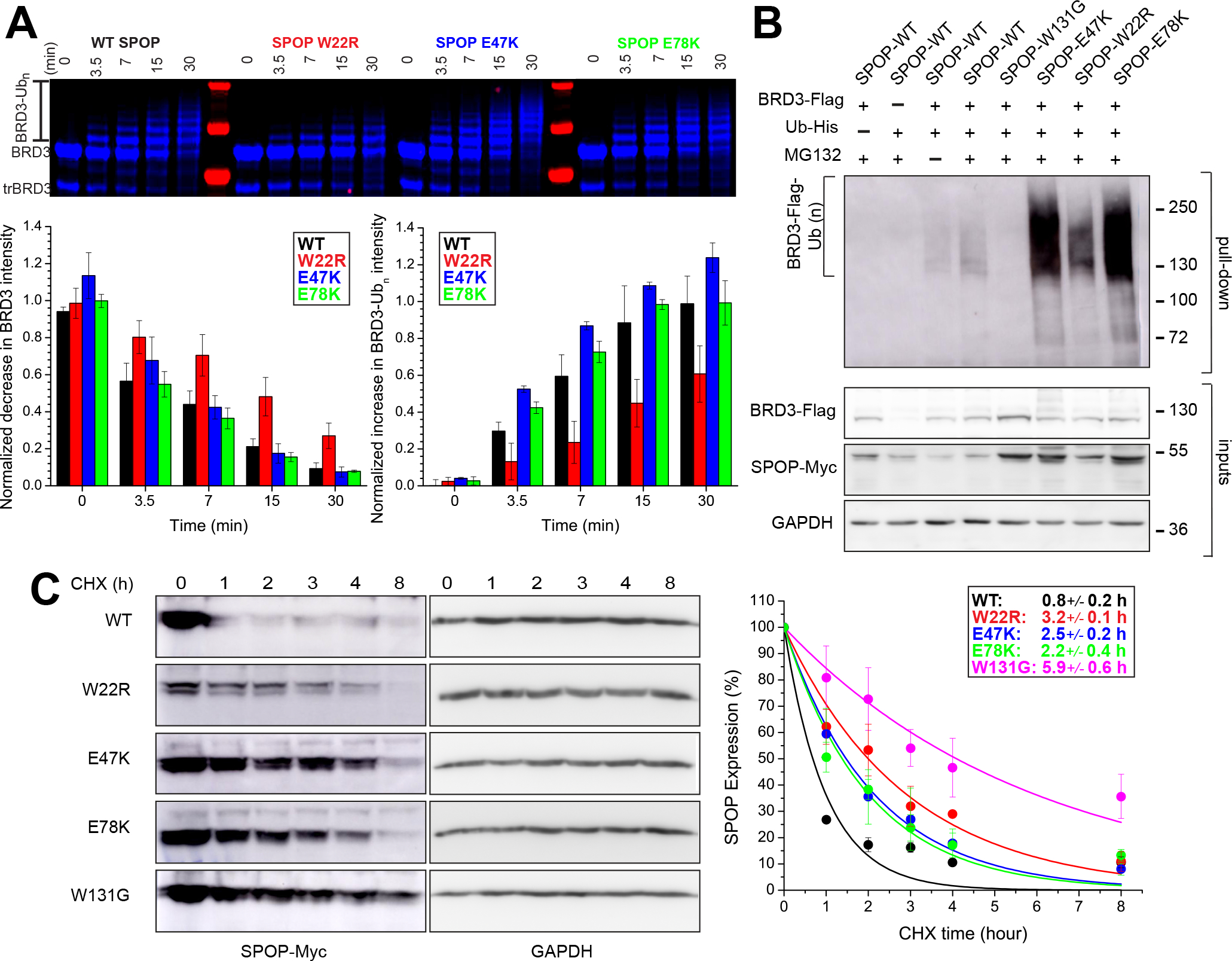
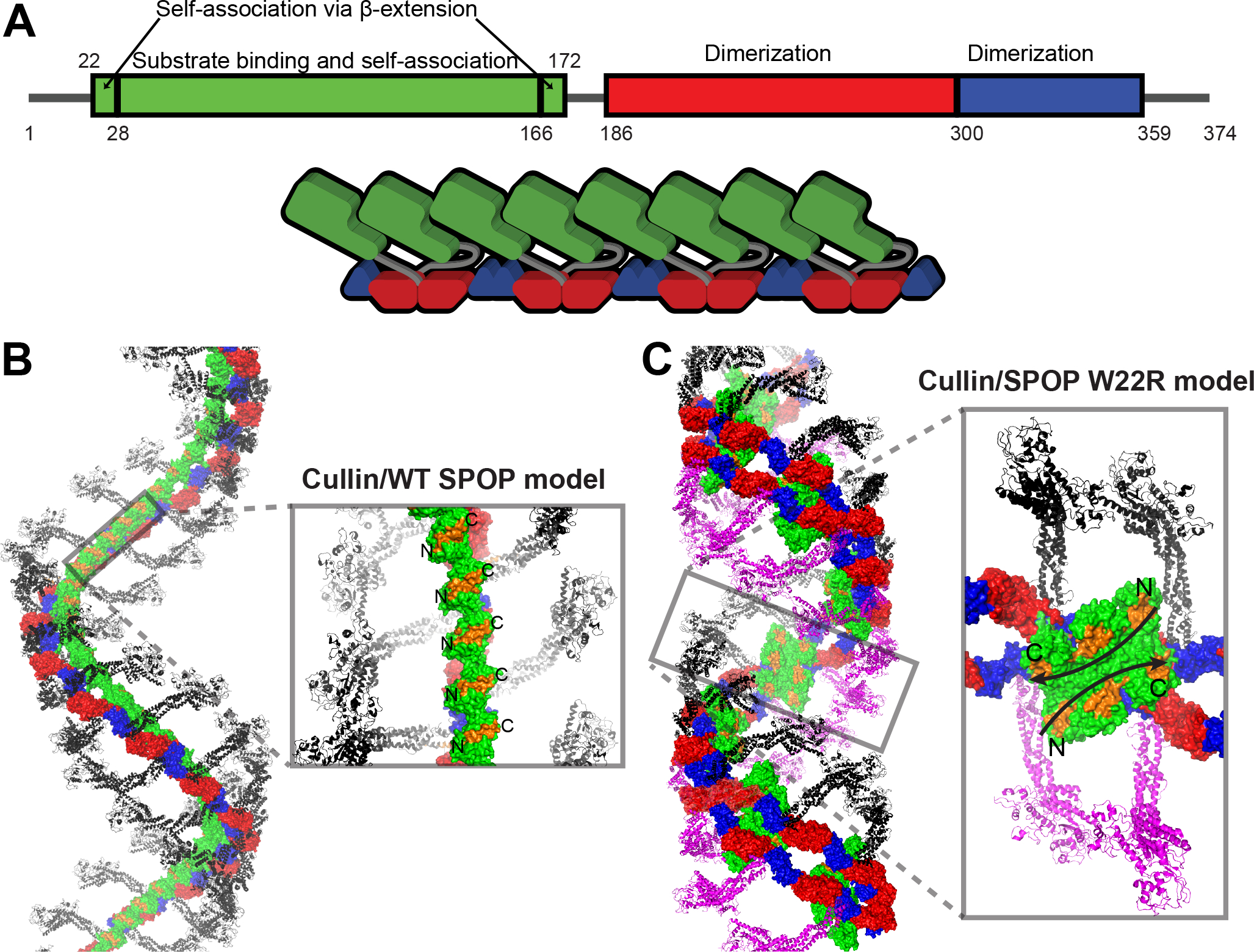
The speckle-type POZ protein (SPOP) functions in the Cullin3-RING ubiquitin ligase (CRL3) as a receptor for the recognition of substrates involved in cell growth, survival, and signaling. SPOP mutations have been attributed to the development of many types of cancers, including prostate and endometrial cancers. Prostate cancer mutations localize in the substrate-binding site of the substrate recognition (MATH) domain and reduce or prevent binding. However, most endometrial cancer mutations are dispersed in seemingly inconspicuous solvent-exposed regions of SPOP, offering no clear basis for their cancer-causing and peculiar gain-of-function properties. Herein, we present the first structure of SPOP in its oligomeric form, uncovering several new interfaces important for SPOP self-assembly and normal function. Given that many previously unaccounted-for cancer mutations are localized in these newly identified interfaces, we uncover molecular mechanisms underlying dysregulation of SPOP function, with effects ranging from gross structural changes to enhanced self-association, and heightened stability and activity.
Keywords: cryo-EM; endometrial cancer; filament; gain of function; higher-order oligomer; hypermorphic; oncogene; prostate cancer; structure; tumor suppressor.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests T.M. was a consultant for Faze Medicines, Inc. and is a member of the journal’s advisory board.
Figures
Comment in
-
Hidden tricks in MATH: Hypermorphic mutations in SPOP tumor suppressor explained by cryo-EM.Mol Cell. 2023 Mar 2;83(5):655-656. doi: 10.1016/j.molcel.2023.02.003. Mol Cell. 2023. PMID: 36868187 Free PMC article.
Similar articles
-
Reduction of oligomer size modulates the competition between cluster formation and phase separation of the tumor suppressor SPOP.J Biol Chem. 2023 Dec;299(12):105427. doi: 10.1016/j.jbc.2023.105427. Epub 2023 Nov 4. J Biol Chem. 2023. PMID: 37926283 Free PMC article.
-
Opposing effects of cancer-type-specific SPOP mutants on BET protein degradation and sensitivity to BET inhibitors.Nat Med. 2017 Sep;23(9):1046-1054. doi: 10.1038/nm.4372. Epub 2017 Aug 14. Nat Med. 2017. PMID: 28805821 Free PMC article.
-
Destruction of DDIT3/CHOP protein by wild-type SPOP but not prostate cancer-associated mutants.Hum Mutat. 2014 Sep;35(9):1142-51. doi: 10.1002/humu.22614. Epub 2014 Jul 23. Hum Mutat. 2014. PMID: 24990631
-
The ubiquitin ligase adaptor SPOP in cancer.FEBS J. 2019 Oct;286(20):3946-3958. doi: 10.1111/febs.15056. Epub 2019 Sep 18. FEBS J. 2019. PMID: 31495053 Free PMC article. Review.
-
The emerging role of speckle-type POZ protein (SPOP) in cancer development.Drug Discov Today. 2014 Sep;19(9):1498-502. doi: 10.1016/j.drudis.2014.07.009. Epub 2014 Jul 21. Drug Discov Today. 2014. PMID: 25058385 Free PMC article. Review.
Cited by
-
SPOP point mutations regulate substrate preference and affect its function.Cell Death Dis. 2024 Feb 26;15(2):172. doi: 10.1038/s41419-024-06565-1. Cell Death Dis. 2024. PMID: 38409107 Free PMC article.
-
The equilibrium between two quaternary assembly states determines the activity of SPOP and its cancer mutants.bioRxiv [Preprint]. 2025 Jun 20:2025.06.19.659812. doi: 10.1101/2025.06.19.659812. bioRxiv. 2025. PMID: 40666954 Free PMC article. Preprint.
-
Cellular Function of a Biomolecular Condensate Is Determined by Its Ultrastructure.bioRxiv [Preprint]. 2024 Dec 27:2024.12.27.630454. doi: 10.1101/2024.12.27.630454. bioRxiv. 2024. PMID: 39763716 Free PMC article. Preprint.
-
Modulating Phase Behavior in Fatty Acid-Modified Elastin-like Polypeptides (FAMEs): Insights into the Impact of Lipid Length on Thermodynamics and Kinetics of Phase Separation.J Am Chem Soc. 2024 Feb 28;146(8):5383-5392. doi: 10.1021/jacs.3c12791. Epub 2024 Feb 14. J Am Chem Soc. 2024. PMID: 38353994 Free PMC article.
-
Hidden tricks in MATH: Hypermorphic mutations in SPOP tumor suppressor explained by cryo-EM.Mol Cell. 2023 Mar 2;83(5):655-656. doi: 10.1016/j.molcel.2023.02.003. Mol Cell. 2023. PMID: 36868187 Free PMC article.
References
-
- Geng C, He B, Xu L, Barbieri CE, Eedunuri VK, Chew SA, Zimmermann M, Bond R, Shou J, Li C, et al. (2013). Prostate cancer-associated mutations in speckle-type POZ protein (SPOP) regulate steroid receptor coactivator 3 protein turnover. Proc Natl Acad Sci U S A 110, 6997–7002. 10.1073/pnas.1304502110. - DOI - PMC - PubMed
-
- Janouskova H, El Tekle G, Bellini E, Udeshi ND, Rinaldi A, Ulbricht A, Bernasocchi T, Civenni G, Losa M, Svinkina T, et al. (2017). Opposing effects of cancer-type-specific SPOP mutants on BET protein degradation and sensitivity to BET inhibitors. Nat Med 23, 1046–1054. 10.1038/nm.4372. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
