

Malaria-driven expansion of adaptive-like functional CD56-negative NK cells correlates with clinical immunity to malaria
- PMID: 36696483
- PMCID: PMC9976268
- DOI: 10.1126/scitranslmed.add9012
Malaria-driven expansion of adaptive-like functional CD56-negative NK cells correlates with clinical immunity to malaria
Abstract
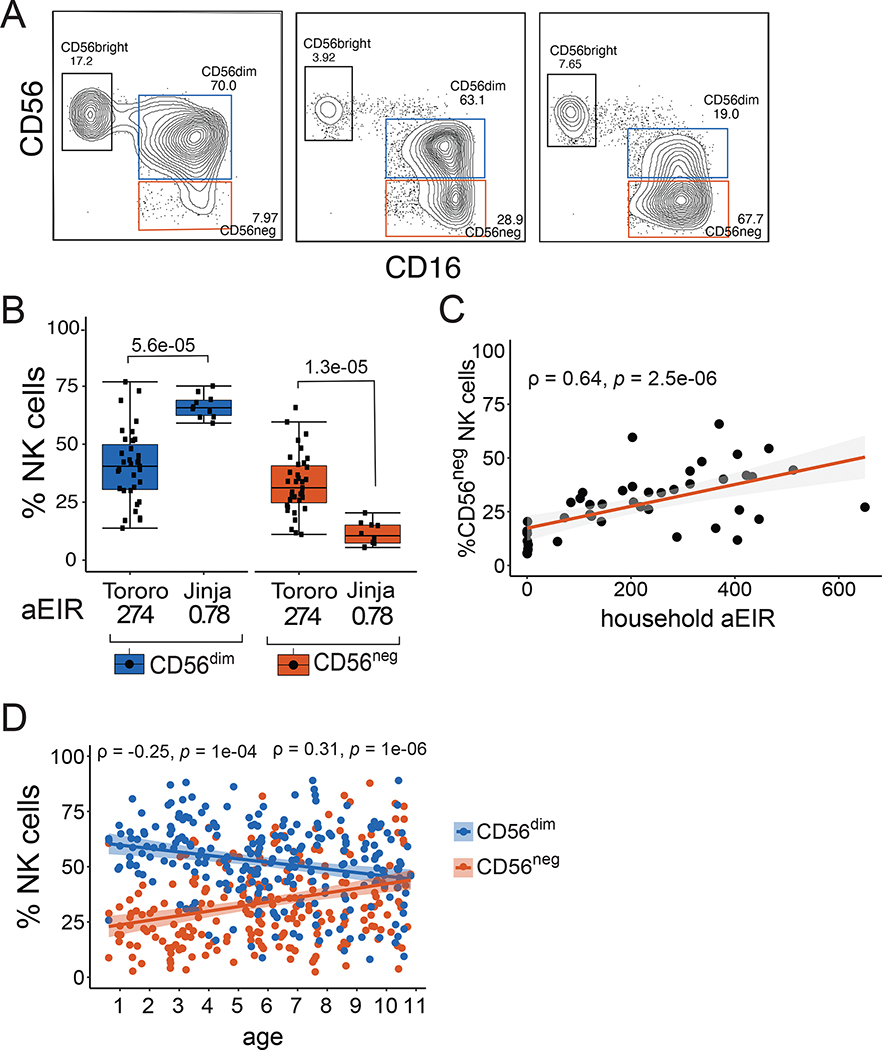
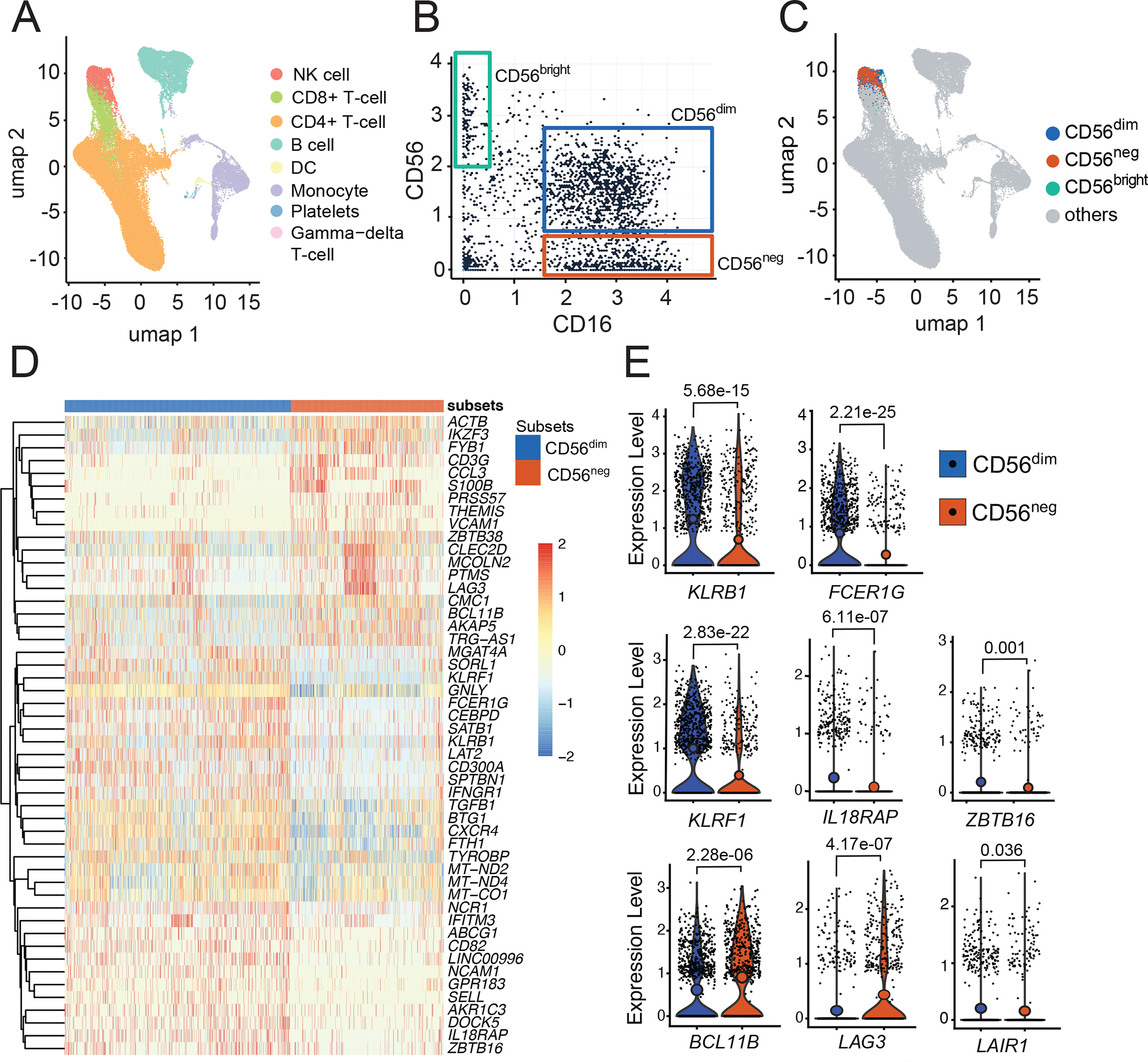
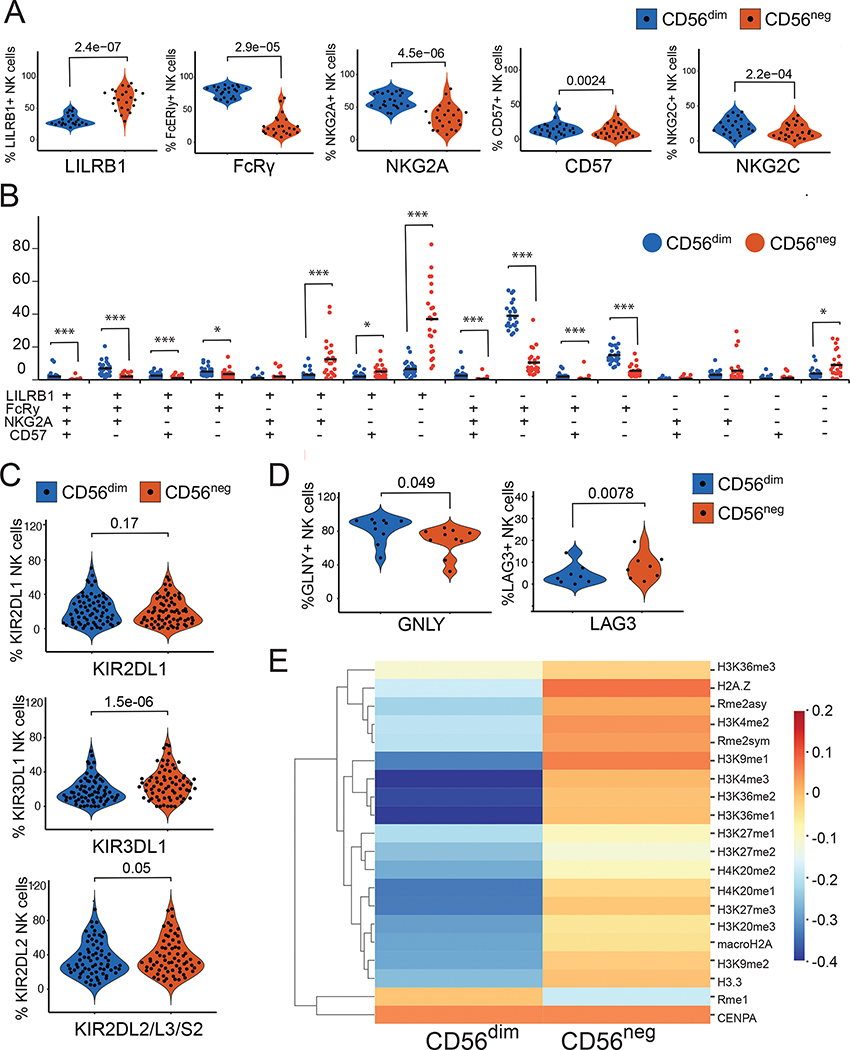
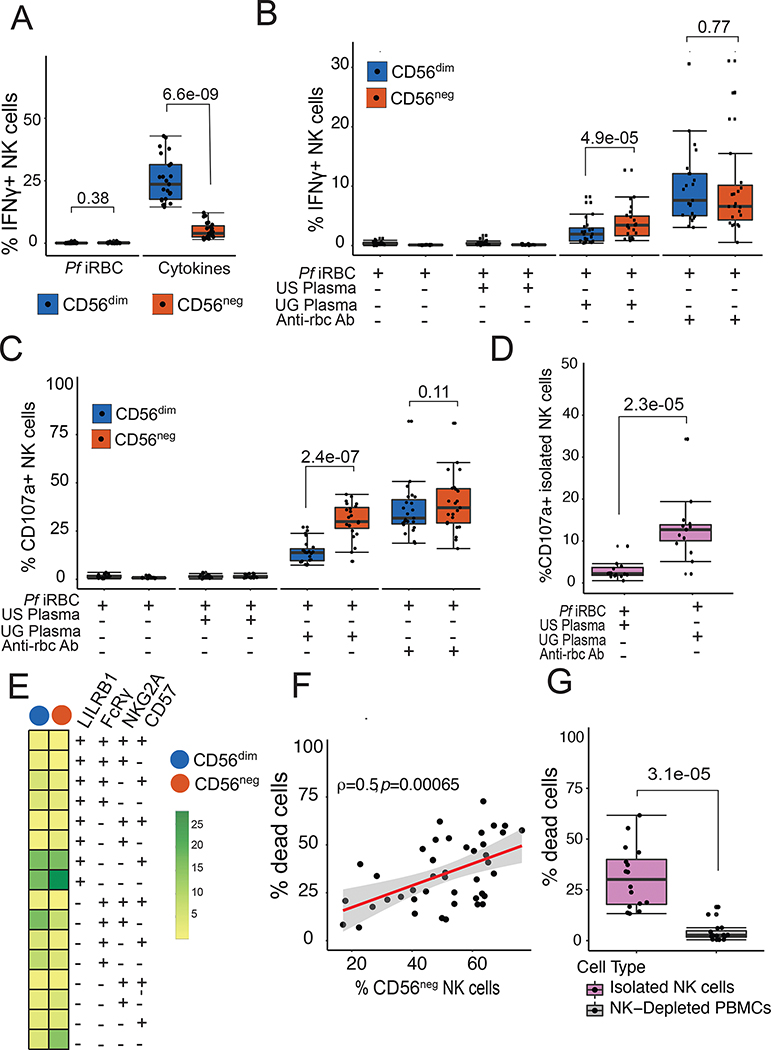
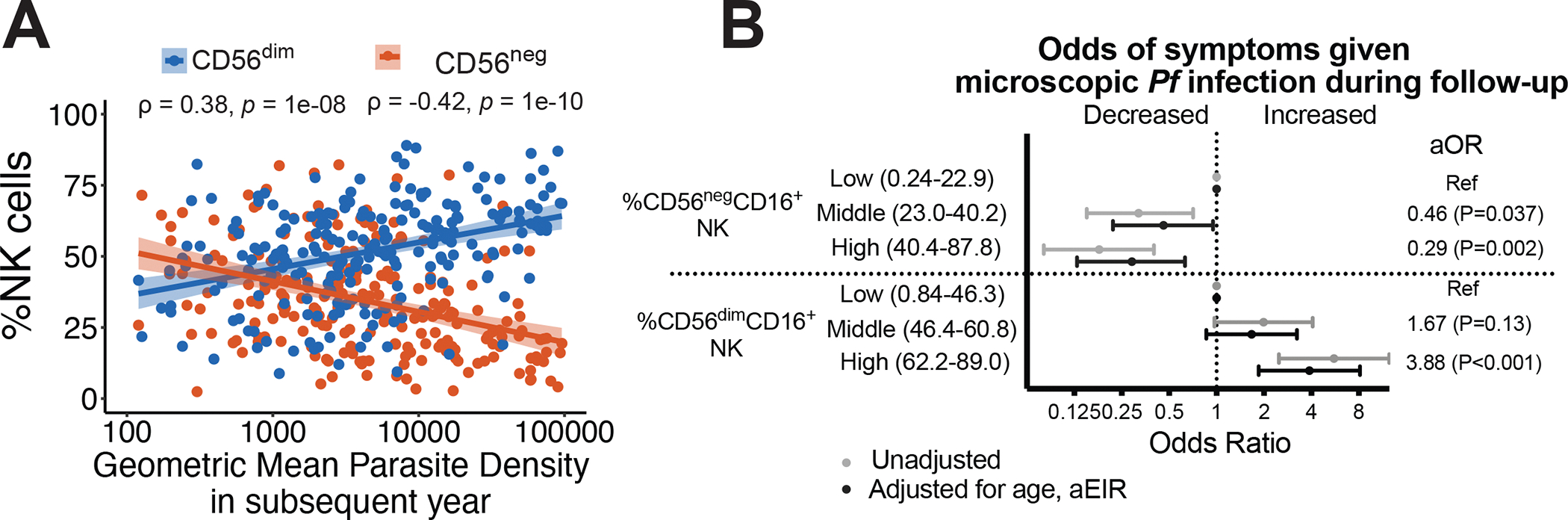
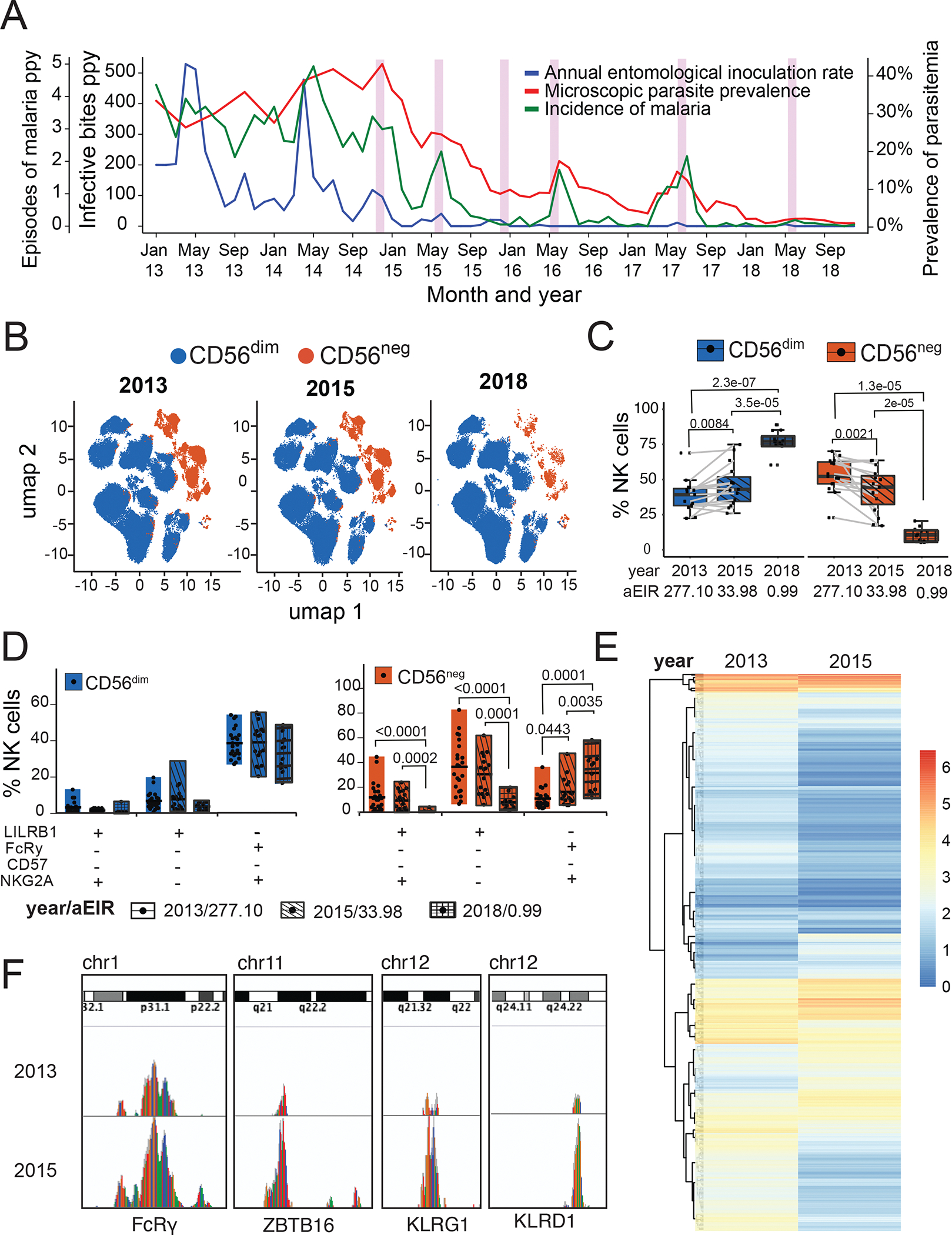
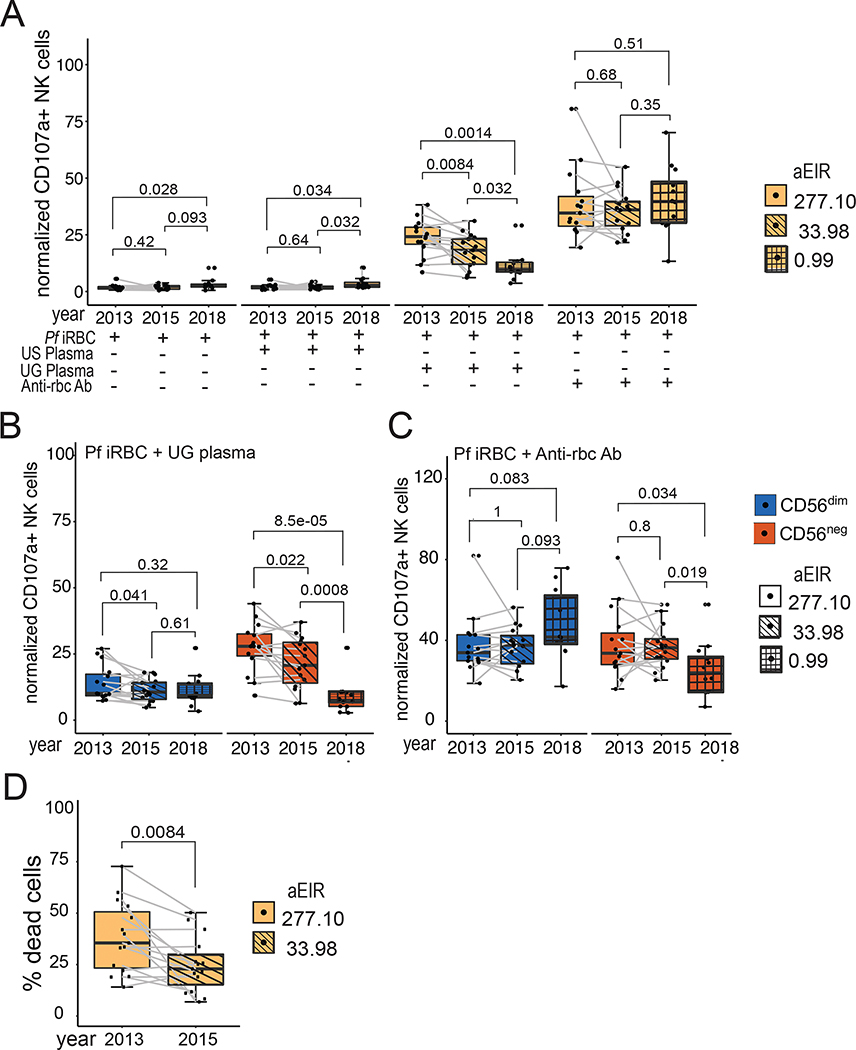
Natural killer (NK) cells likely play an important role in immunity to malaria, but the effect of repeated malaria on NK cell responses remains unclear. Here, we comprehensively profiled the NK cell response in a cohort of 264 Ugandan children. Repeated malaria exposure was associated with expansion of an atypical, CD56neg population of NK cells that differed transcriptionally, epigenetically, and phenotypically from CD56dim NK cells, including decreased expression of PLZF and the Fc receptor γ-chain, increased histone methylation, and increased protein expression of LAG-3, KIR, and LILRB1. CD56neg NK cells were highly functional and displayed greater antibody-dependent cellular cytotoxicity than CD56dim NK cells. Higher frequencies of CD56neg NK cells were associated with protection against symptomatic malaria and high parasite densities. After marked reductions in malaria transmission, frequencies of these cells rapidly declined, suggesting that continuous exposure to Plasmodium falciparum is required to maintain this modified, adaptive-like NK cell subset.
Conflict of interest statement
Figures
References
-
- “World malaria report 2021,” (World Health Organization, Geneva, Switzerland, 2021).
-
- Rodriguez-Barraquer I, Arinaitwe E, Jagannathan P, Kamya MR, Rosenthal PJ, Rek J, Dorsey G, Nankabirwa J, Staedke SG, Kilama M, Drakeley C, Ssewanyana I, Smith DL, Greenhouse B, Quantification of anti-parasite and anti-disease immunity to malaria as a function of age and exposure. Elife 7, (2018). - PMC - PubMed
-
- Langhorne J, Ndungu FM, Sponaas AM, Marsh K, Immunity to malaria: more questions than answers. Nat Immunol 9, 725–732 (2008). - PubMed
-
- Tan J, Piccoli L, Lanzavecchia A, The Antibody Response to Plasmodium falciparum: Cues for Vaccine Design and the Discovery of Receptor-Based Antibodies. Annu Rev Immunol, (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
