

Brainstem astrocytes control homeostatic regulation of caloric intake
- PMID: 36696965
- PMCID: PMC10026361
- DOI: 10.1113/JP283566
Brainstem astrocytes control homeostatic regulation of caloric intake
Abstract
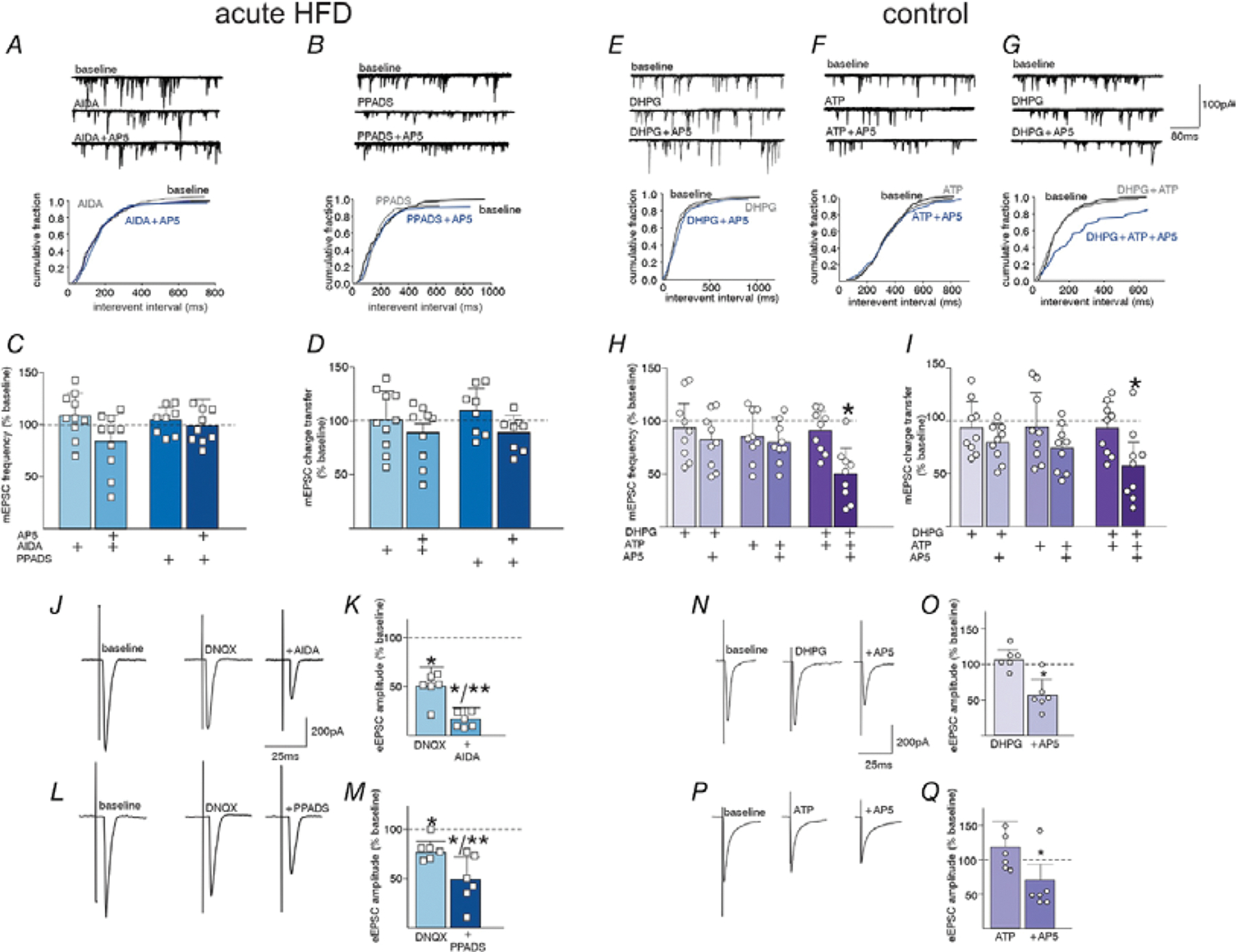
Prolonged high-fat diet (HFD) exposure is associated with hyperphagia, excess caloric intake and weight gain. After initial exposure to a HFD, a brief (24-48 h) period of hyperphagia is followed by the regulation of caloric intake and restoration of energy balance within an acute (3-5 day) period. Previous studies have demonstrated this occurs via a vagally mediated signalling cascade that increases glutamatergic transmission via activation of NMDA receptors located on gastric-projecting neurons of the dorsal motor nucleus of the vagus (DMV). The present study used electrophysiological recordings from thin brainstem slice preparations, in vivo recordings of gastric motility and tone, measurement of gastric emptying rates, and food intake studies to investigate the hypothesis that activation of brainstem astrocytes in response to acute HFD exposure is responsible for the increased glutamatergic drive to DMV neurons and the restoration of caloric balance. Pharmacological and chemogenetic inhibition of brainstem astrocytes reduced glutamatergic signalling and DMV excitability, dysregulated gastric tone and motility, attenuated the homeostatic delay in gastric emptying, and prevented the decrease in food intake that is observed during the period of energy regulation following initial exposure to HFD. Understanding the mechanisms involved in caloric regulation may provide critical insights into energy balance as well as into the hyperphagia that develops as these mechanisms are overcome. KEY POINTS: Initial exposure to a high fat diet is associated with a brief period of hyperphagia before caloric intake and energy balance is restored. This period of homeostatic regulation is associated with a vagally mediated signalling cascade that increases glutamatergic transmission to dorsal motor nucleus of the vagus (DMV) neurons via activation of synaptic NMDA receptors. The present study demonstrates that pharmacological and chemogenetic inhibition of brainstem astrocytes reduced glutamatergic signalling and DMV neuronal excitability, dysregulated gastric motility and tone and emptying, and prevented the regulation of food intake following high-fat diet exposure. Astrocyte regulation of glutamatergic transmission to DMV neurons appears to involve release of the gliotransmitters glutamate and ATP. Understanding the mechanisms involved in caloric regulation may provide critical insights into energy balance as well as into the hyperphagia that develops as these mechanisms are overcome.
Keywords: NMDA receptor; astrocyte; feeding; gastroenterology; high fat diet; neurophysiology; neuroplasticity; obesity.
© 2022 The Authors. The Journal of Physiology © 2022 The Physiological Society.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests
Figures








Comment in
-
Gastrocytes and GLUttony - astrocyte regulation of calorie intake via glutamatergic modulation of gastric activity in rats.J Physiol. 2023 Feb;601(4):709-710. doi: 10.1113/JP284264. Epub 2023 Feb 2. J Physiol. 2023. PMID: 36704958 No abstract available.
Similar articles
-
Acute high-fat diet upregulates glutamatergic signaling in the dorsal motor nucleus of the vagus.Am J Physiol Gastrointest Liver Physiol. 2018 May 1;314(5):G623-G634. doi: 10.1152/ajpgi.00395.2017. Epub 2018 Jan 25. Am J Physiol Gastrointest Liver Physiol. 2018. PMID: 29368945 Free PMC article.
-
DMV extrasynaptic NMDA receptors regulate caloric intake in rats.JCI Insight. 2021 May 10;6(9):e139785. doi: 10.1172/jci.insight.139785. JCI Insight. 2021. PMID: 33764905 Free PMC article.
-
Exposure to a high fat diet during the perinatal period alters vagal motoneurone excitability, even in the absence of obesity.J Physiol. 2015 Jan 1;593(1):285-303. doi: 10.1113/jphysiol.2014.282806. Epub 2014 Dec 2. J Physiol. 2015. PMID: 25556801 Free PMC article.
-
Glutamatergic plasticity within neurocircuits of the dorsal vagal complex and the regulation of gastric functions.Am J Physiol Gastrointest Liver Physiol. 2021 May 1;320(5):G880-G887. doi: 10.1152/ajpgi.00014.2021. Epub 2021 Mar 17. Am J Physiol Gastrointest Liver Physiol. 2021. PMID: 33730858 Free PMC article. Review.
-
Fatty acid-induced astrocyte ketone production and the control of food intake.Am J Physiol Regul Integr Comp Physiol. 2016 Jun 1;310(11):R1186-92. doi: 10.1152/ajpregu.00113.2016. Epub 2016 Apr 27. Am J Physiol Regul Integr Comp Physiol. 2016. PMID: 27122369 Free PMC article. Review.
Cited by
-
Constant light and high fat diet alter daily patterns of activity, feed intake and fecal corticosterone levels in pregnant and lactating female ICR mice.PLoS One. 2024 Nov 20;19(11):e0312419. doi: 10.1371/journal.pone.0312419. eCollection 2024. PLoS One. 2024. PMID: 39565751 Free PMC article.
-
Synaptic plasticity and the role of astrocytes in central metabolic circuits.WIREs Mech Dis. 2024 Jan-Feb;16(1):e1632. doi: 10.1002/wsbm.1632. Epub 2023 Oct 13. WIREs Mech Dis. 2024. PMID: 37833830 Free PMC article. Review.
-
GABAAR-δ-subunit mediates increased GABAergic inhibition in cardiac DMV neurons after high-fat diet.iScience. 2025 Mar 22;28(4):112268. doi: 10.1016/j.isci.2025.112268. eCollection 2025 Apr 18. iScience. 2025. PMID: 40264791 Free PMC article.
-
Sex differences in central endozepine expression and regulation of appetite.Physiol Behav. 2025 Oct 1;299:115003. doi: 10.1016/j.physbeh.2025.115003. Epub 2025 Jun 17. Physiol Behav. 2025. PMID: 40553808
-
Gastrointestinal dysfunction in the valproic acid induced model of social deficit in rats.Auton Neurosci. 2024 Jun;253:103161. doi: 10.1016/j.autneu.2024.103161. Epub 2024 Feb 29. Auton Neurosci. 2024. PMID: 38461695 Free PMC article.
References
-
- Accorsi-Mendonca D, Bonagamba LGH & Machado BH. (2019). Astrocytic modulation of glutamatergic synaptic transmission is reduced in NTS of rats submitted to short-term sustained hypoxia. J Neurophysiol 121, 1822–1830. - PubMed
-
- Astiz M, Pernia O, Barrios V, Garcia-Segura LM & Diz-Chaves Y. (2017). Short-Term High-Fat Diet Feeding Provides Hypothalamic but Not Hippocampal Protection against Acute Infection in Male Mice. Neuroendocrinology 104, 40–50. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources