

Epigenetic and transcriptional regulations prime cell fate before division during human pluripotent stem cell differentiation
- PMID: 36697417
- PMCID: PMC9876972
- DOI: 10.1038/s41467-023-36116-9
Epigenetic and transcriptional regulations prime cell fate before division during human pluripotent stem cell differentiation
Abstract
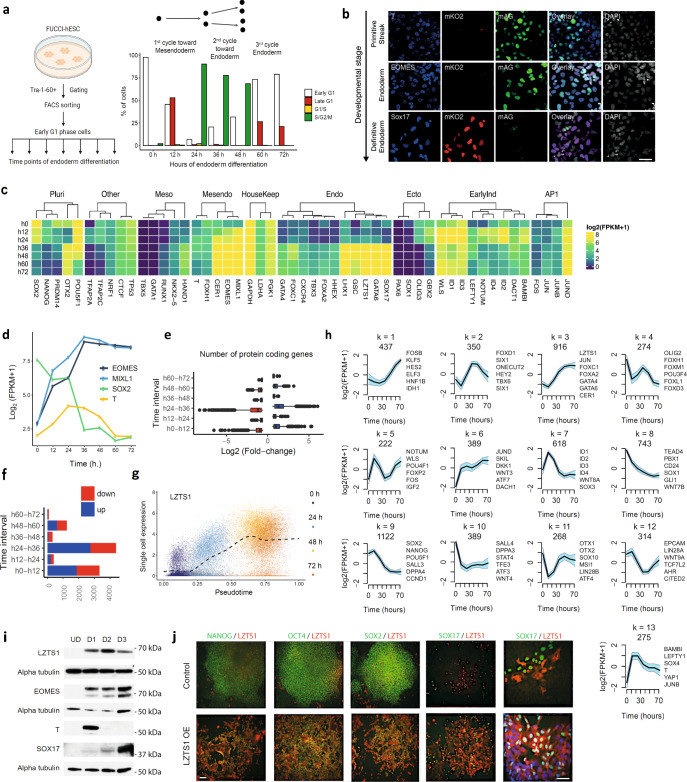
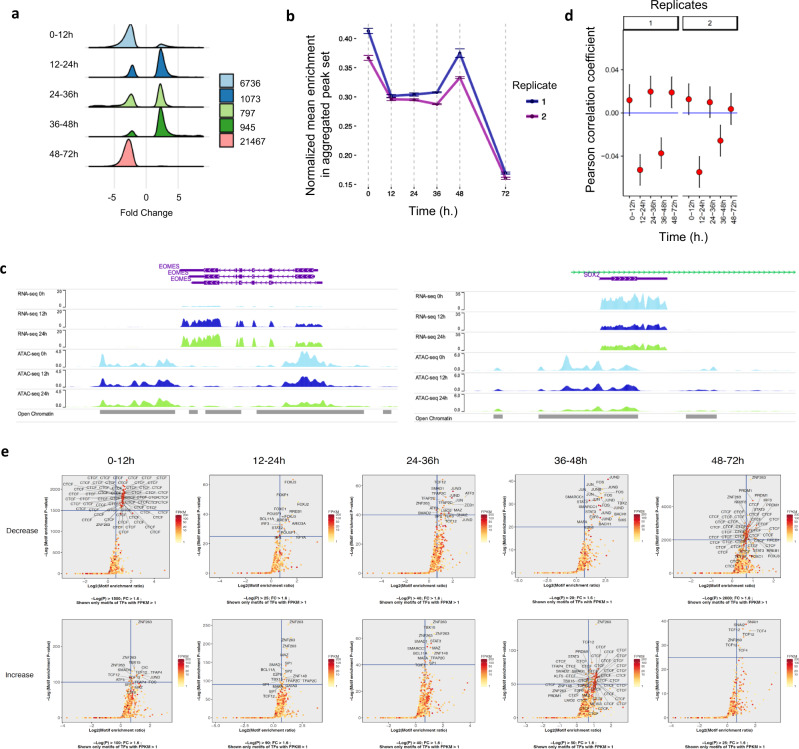
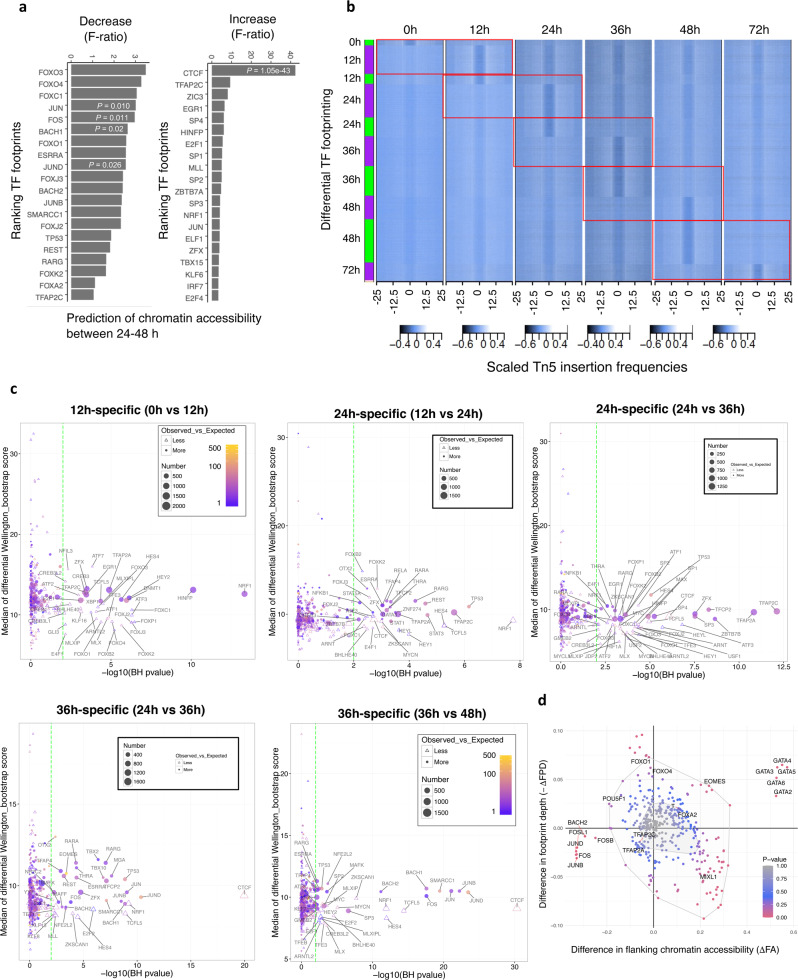
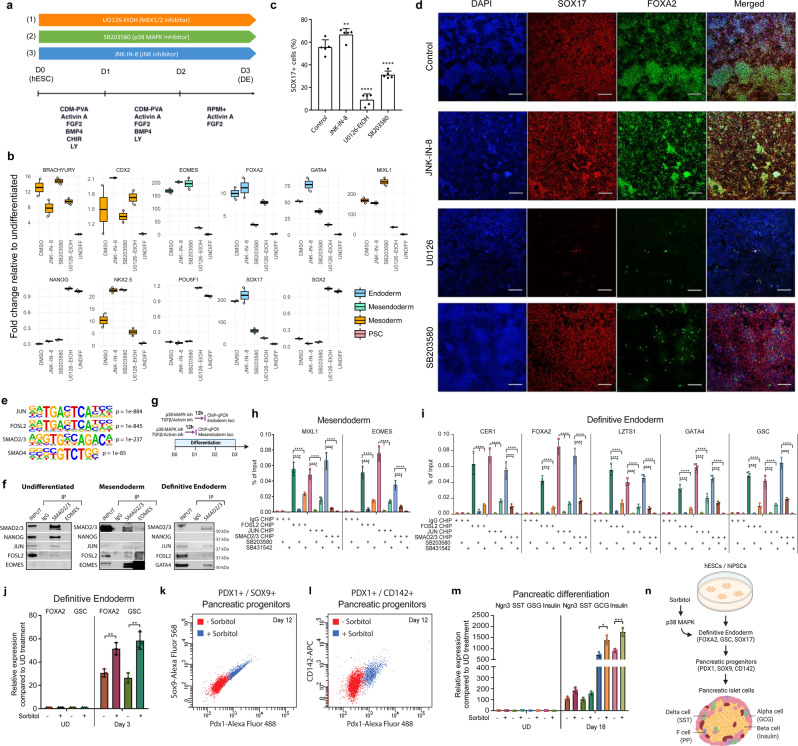
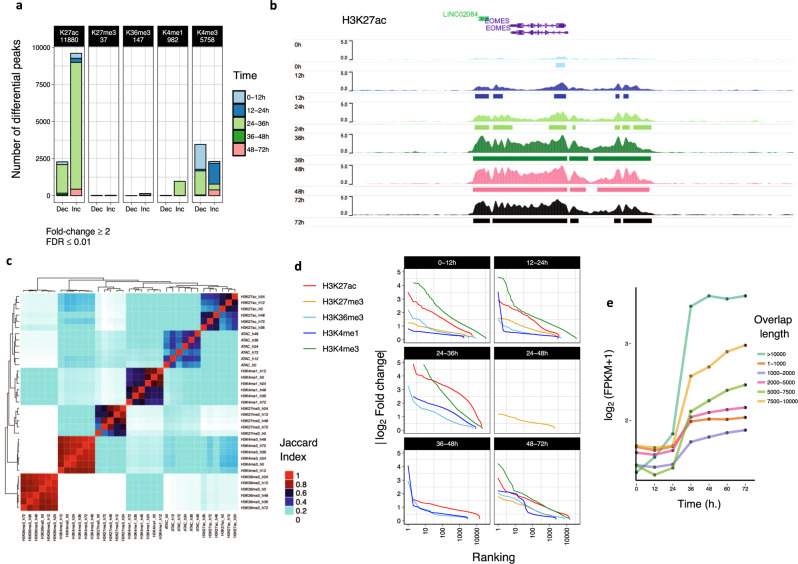
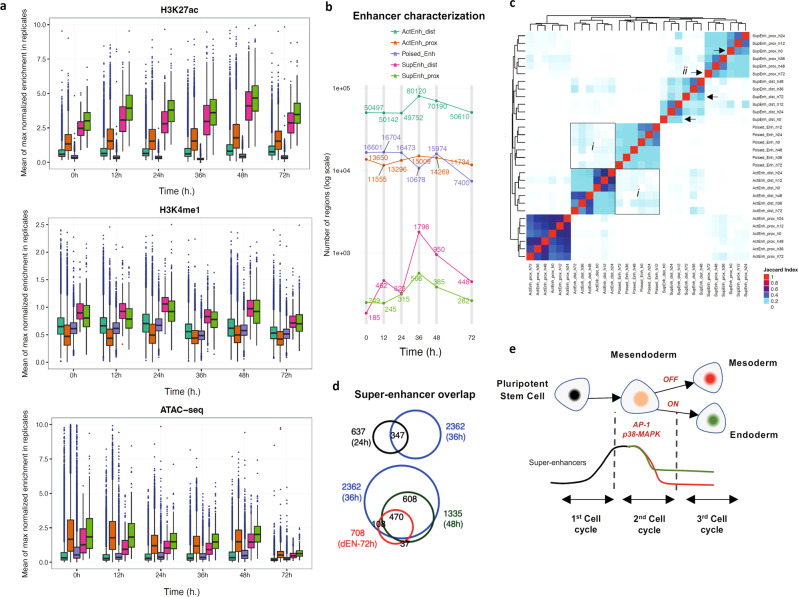
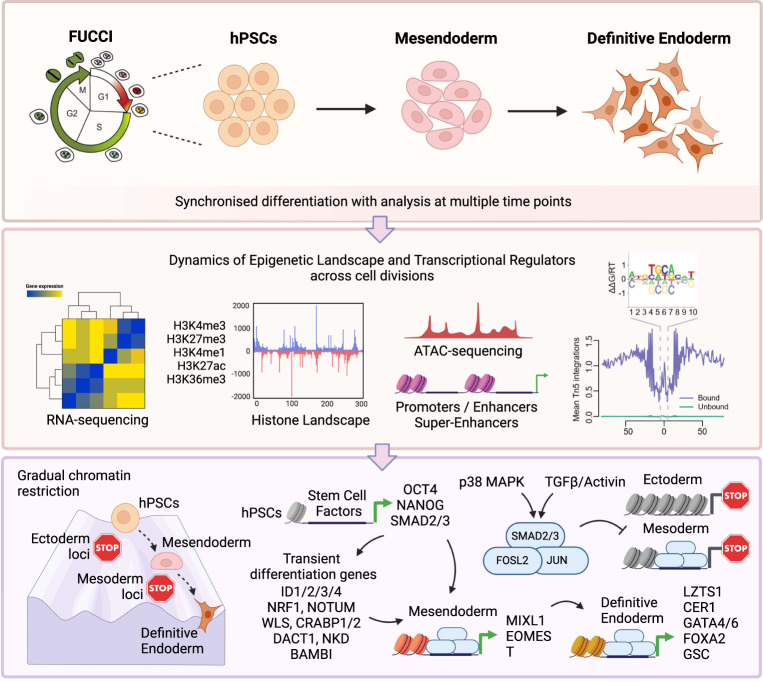
Stem cells undergo cellular division during their differentiation to produce daughter cells with a new cellular identity. However, the epigenetic events and molecular mechanisms occurring between consecutive cell divisions have been insufficiently studied due to technical limitations. Here, using the FUCCI reporter we developed a cell-cycle synchronised human pluripotent stem cell (hPSC) differentiation system for uncovering epigenome and transcriptome dynamics during the first two divisions leading to definitive endoderm. We observed that transcription of key differentiation markers occurs before cell division, while chromatin accessibility analyses revealed the early inhibition of alternative cell fates. We found that Activator protein-1 members controlled by p38/MAPK signalling are necessary for inducing endoderm while blocking cell fate shifting toward mesoderm, and that enhancers are rapidly established and decommissioned between different cell divisions. Our study has practical biomedical utility for producing hPSC-derived patient-specific cell types since p38/MAPK induction increased the differentiation efficiency of insulin-producing pancreatic beta-cells.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Efficient endoderm induction from human pluripotent stem cells by logically directing signals controlling lineage bifurcations.Cell Stem Cell. 2014 Feb 6;14(2):237-52. doi: 10.1016/j.stem.2013.12.007. Epub 2014 Jan 9. Cell Stem Cell. 2014. PMID: 24412311 Free PMC article.
-
Cell-context response to germ layer differentiation signals is predetermined by the epigenome in regionalized epiblast populations.Nat Commun. 2025 May 29;16(1):5000. doi: 10.1038/s41467-025-60348-6. Nat Commun. 2025. PMID: 40442089 Free PMC article.
-
Generation of vascularized pancreatic progenitors through co-differentiation of endoderm and mesoderm from human pluripotent stem cells.Stem Cell Res Ther. 2024 Dec 24;15(1):502. doi: 10.1186/s13287-024-04120-5. Stem Cell Res Ther. 2024. PMID: 39719603 Free PMC article.
-
Human parthenogenetic stem cells produce enriched populations of definitive endoderm cells after trichostatin A pretreatment.Differentiation. 2011 Jun;81(5):292-8. doi: 10.1016/j.diff.2011.01.002. Epub 2011 Feb 8. Differentiation. 2011. PMID: 21306817 Review.
-
Metabolic and epigenetic regulation of endoderm differentiation.Trends Cell Biol. 2022 Feb;32(2):151-164. doi: 10.1016/j.tcb.2021.09.002. Epub 2021 Oct 1. Trends Cell Biol. 2022. PMID: 34607773 Free PMC article. Review.
Cited by
-
CTCF deletion alters the pluripotency and DNA methylation profile of human iPSCs.Front Cell Dev Biol. 2023 Nov 30;11:1302448. doi: 10.3389/fcell.2023.1302448. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 38099298 Free PMC article.
-
The epigenetic landscape in intestinal stem cells and its deregulation in colorectal cancer.Stem Cells. 2024 Jun 14;42(6):509-525. doi: 10.1093/stmcls/sxae027. Stem Cells. 2024. PMID: 38597726 Free PMC article. Review.
-
Possible Strategies to Reduce the Tumorigenic Risk of Reprogrammed Normal and Cancer Cells.Int J Mol Sci. 2024 May 9;25(10):5177. doi: 10.3390/ijms25105177. Int J Mol Sci. 2024. PMID: 38791215 Free PMC article. Review.
-
SMAD2/3-SMYD2 and developmental transcription factors cooperate with cell-cycle inhibitors to guide tissue formation.Protein Cell. 2025 Apr 18;16(4):260-285. doi: 10.1093/procel/pwae031. Protein Cell. 2025. PMID: 38758030 Free PMC article.
-
Time-dependent changes in genome-wide gene expression and post-transcriptional regulation across the post-death process in silkworm.DNA Res. 2024 Dec 1;31(6):dsae031. doi: 10.1093/dnares/dsae031. DNA Res. 2024. PMID: 39546332 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
