

Acyl-chain saturation regulates the order of phosphatidylinositol 4,5-bisphosphate nanodomains
- PMID: 36697613
- PMCID: PMC9814227
- DOI: 10.1038/s42004-021-00603-1
Acyl-chain saturation regulates the order of phosphatidylinositol 4,5-bisphosphate nanodomains
Abstract
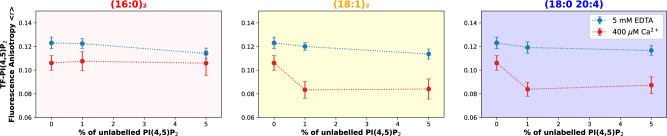
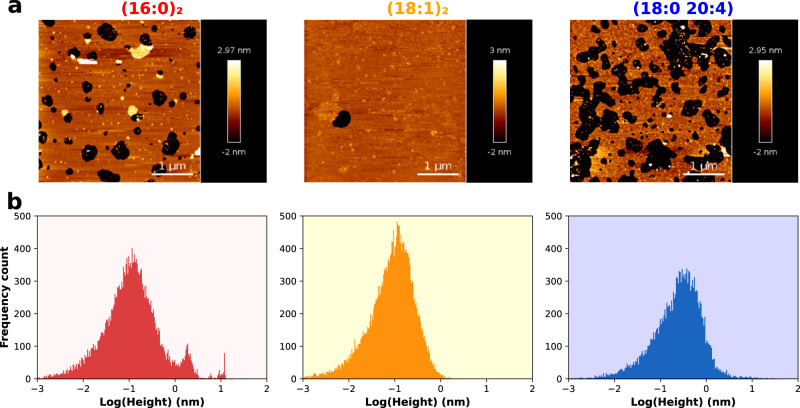
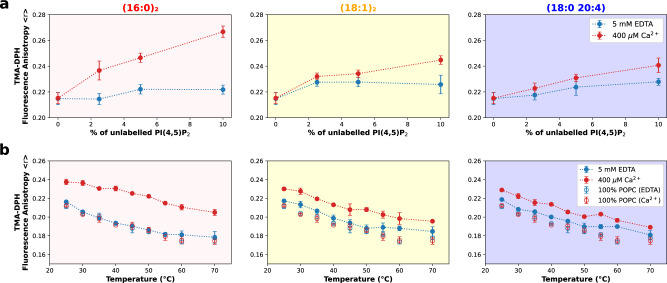
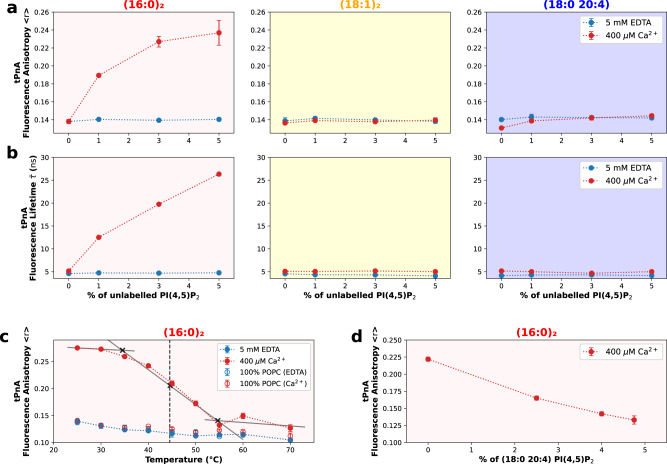
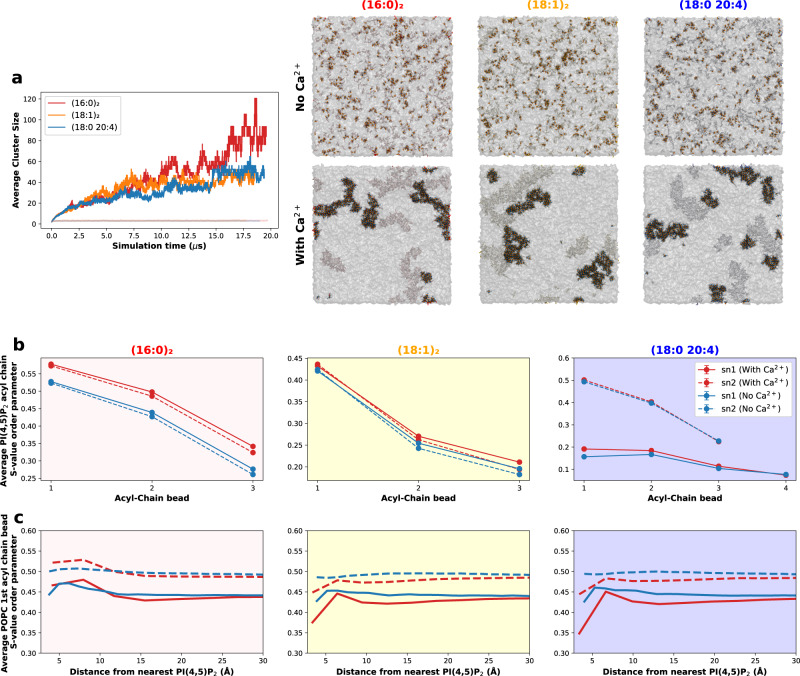
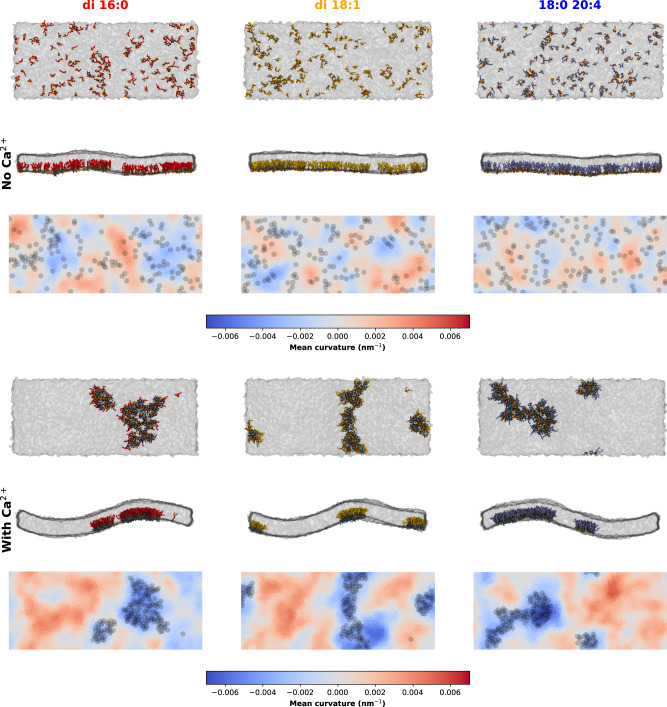
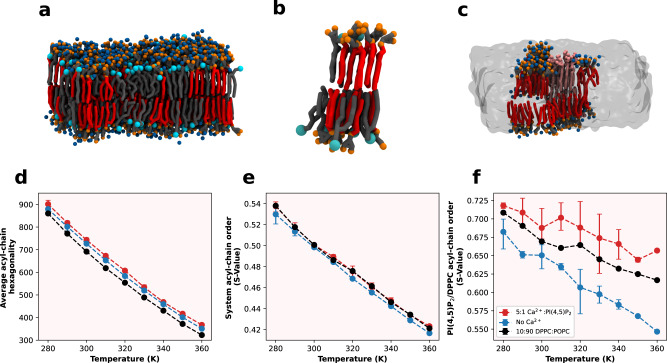
Phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2) plays a critical role in the regulation of various plasma membrane processes and signaling pathways in eukaryotes. A significant amount of cellular resources are spent on maintaining the dominant 1-stearoyl-2-arachidonyl PI(4,5)P2 acyl-chain composition, while less abundant and more saturated species become more prevalent in response to specific stimuli, stress or aging. Here, we report the impact of acyl-chain structure on the biophysical properties of cation-induced PI(4,5)P2 nanodomains. PI(4,5)P2 species with increasing levels of acyl-chain saturation cluster in progressively more ordered nanodomains, culminating in the formation of gel-like nanodomains for fully saturated species. The formation of these gel-like domains was largely abrogated in the presence of 1-stearoyl-2-arachidonyl PI(4,5)P2. This is, to the best of our knowledge, the first report of the impact of PI(4,5)P2 acyl-chain composition on cation-dependent nanodomain ordering, and provides important clues to the motives behind the enrichment of PI(4,5)P2 with polyunsaturated acyl-chains. We also show how Ca2+-induced PI(4,5)P2 nanodomains are able to generate local negative curvature, a phenomenon likely to play a role in membrane remodeling events.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Impact of Ca2+-Induced PI(4,5)P2 Clusters on PH-YFP Organization and Protein-Protein Interactions.Biomolecules. 2022 Jun 29;12(7):912. doi: 10.3390/biom12070912. Biomolecules. 2022. PMID: 35883468 Free PMC article.
-
Phosphatidylinositol-(4,5)-Bisphosphate Acyl Chains Differentiate Membrane Binding of HIV-1 Gag from That of the Phospholipase Cδ1 Pleckstrin Homology Domain.J Virol. 2015 Aug;89(15):7861-73. doi: 10.1128/JVI.00794-15. Epub 2015 May 20. J Virol. 2015. PMID: 25995263 Free PMC article.
-
The acyltransferase LYCAT controls specific phosphoinositides and related membrane traffic.Mol Biol Cell. 2017 Jan 1;28(1):161-172. doi: 10.1091/mbc.E16-09-0668. Epub 2016 Nov 9. Mol Biol Cell. 2017. PMID: 28035047 Free PMC article.
-
Enrichment of phosphatidylinositols with specific acyl chains.Biochim Biophys Acta. 2014 Jun;1838(6):1501-8. doi: 10.1016/j.bbamem.2013.10.003. Epub 2013 Oct 10. Biochim Biophys Acta. 2014. PMID: 24120446 Review.
-
Structure and Lateral Organization of Phosphatidylinositol 4,5-bisphosphate.Molecules. 2020 Aug 26;25(17):3885. doi: 10.3390/molecules25173885. Molecules. 2020. PMID: 32858905 Free PMC article. Review.
Cited by
-
Effects of Ca2+ on the Structure and Dynamics of PIP3 in Model Membranes Containing PC and PS.Biochemistry. 2025 Jan 7;64(1):127-137. doi: 10.1021/acs.biochem.4c00513. Epub 2024 Dec 10. Biochemistry. 2025. PMID: 39656263
-
Impact of Ca2+-Induced PI(4,5)P2 Clusters on PH-YFP Organization and Protein-Protein Interactions.Biomolecules. 2022 Jun 29;12(7):912. doi: 10.3390/biom12070912. Biomolecules. 2022. PMID: 35883468 Free PMC article.
-
The Influence of Phosphoinositide Lipids in the Molecular Biology of Membrane Proteins: Recent Insights from Simulations.J Mol Biol. 2025 Feb 15;437(4):168937. doi: 10.1016/j.jmb.2025.168937. Epub 2025 Jan 9. J Mol Biol. 2025. PMID: 39793883 Free PMC article. Review.
-
Effects of cholesterol and PIP2 on interactions between glycophorin A and Band 3 in lipid bilayers.Biophys J. 2022 Jun 7;121(11):2069-2077. doi: 10.1016/j.bpj.2022.05.001. Epub 2022 May 6. Biophys J. 2022. PMID: 35524411 Free PMC article.
References
-
- Monteiro ME, Sarmento MJ, Fernandes F. Role of calcium in membrane interactions by PI(4,5)P2-binding proteins. Biochem. Soc. Trans. 2014;42:1441–6. - PubMed
-
- Di Paolo G, De Camilli P. Phosphoinositides in cell regulation and membrane dynamics. Nature. 2006;443:651–657. - PubMed
-
- Koch M, Holt M. Coupling exo- and endocytosis: an essential role for PIP2 at the synapse. Biochim. Biophys. Acta. 2012;1821:1114–1132. - PubMed
-
- Martin, T. F. J. in Subcellular Biochemistry Vol. 59, 111–130 (Springer New York, 2012). - PubMed
-
- dos Remedios, C. G. & Nosworthy, N. J. in Actin-Binding Proteins and Disease 290–297, 10.1007/978-0-387-71749-4_12 (Springer New York, 2008).
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
